Article Text

Abstract
Skeletal muscles are composed of fibres of different types, each type being identified by the isoform of myosin heavy chain which is expressed as slow 1, fast 2A, fast 2X, and fast 2B. Slow fibres are resistant to fatigue due to their highly oxidative metabolism whereas 2X and 2B fibres are easily fatiguable and fast 2A fibres exhibit intermediate fatigue resistance. Slow fibres and fast fibres are present in equal proportions in the adult human diaphragm while intercostal muscles contain a higher proportion of fast fibres. A small fibre size, abundance of capillaries, and a high aerobic oxidative enzyme activity are typical features of diaphragm fibres and give them the resistance to fatigue required by their continuous activity. Because of their fibre composition, intercostal muscles are less resistant to fatigue. The structural and functional characteristics of respiratory muscle fibres are not fixed, however, and can be modified in response to several physiological and pathological conditions such as training (adaptation to changes in respiratory load), adaptation to hypoxia, age related changes, and changes associated with respiratory diseases. The properties of respiratory muscle fibres can also be modified by pharmacological agents such as β2 agonists and corticosteroids used for the treatment of respiratory diseases.
- respiratory muscles
- myosin isoforms
- diaphragm
Statistics from Altmetric.com
The principal function of the lung is to ventilate the blood. The alternating air flow to and from the alveolar surface is driven by pressure gradients generated by the respiratory muscles. In spite of their specific task which does not allow them to rest during their entire life, the respiratory muscles have the same structure and function as all other limb and trunk muscles. The specialisation of the respiratory muscles derives directly from the characteristics of the fibres of which they are composed. It is well known that skeletal muscle fibres are very heterogeneous: the aim of this review is to identify the features of respiratory muscle fibres which enable them to work without becoming fatigued. Respiratory muscle fibres, however, are not only highly specialised for their functional tasks but are also able to modify their properties to adapt to new requirements which might arise from physiological conditions such as physical exercise or from lung or respiratory diseases. Similarly, changes to the respiratory muscle fibres as a result of muscular disease might severely impair respiratory function.
HETEROGENEITY OF MUSCLE FIBRES
Muscle fibres are not equal: the ability to generate force and mechanical power, the time course of the contractile response, and the resistance to fatigue differ markedly from fibre to fibre. On the basis of their functional properties, fibres can be divided into large groups generally known as “fibre types”.1,2 There are several classifications of fibre types, the most common of which are shown in table 1.
Classification of human skeletal muscle fibres and distinctive functional or molecular properties of each fibre type
Modern molecular biology shows that each fibre type is the result of a specific profile of gene expression based on two main mechanisms:1 (1) quantitative expression regulation (upregulation or downregulation) for many genes; and (2) selective expression for other genes which exist in multiple copies with slight differences (“isogenes”) and give rise to isoforms—that is, proteins similar enough to replace each other but diverse enough to give the fibre distinct functional properties. A recent comparison based on microarray hybridisation techniques on a set of 6519 genes has shown that 177 genes (about 3%) are differentially expressed in slow and fast muscles.3
The ability of a muscle fibre to generate tension, to shorten, and to produce mechanical power is mainly determined by its myofibrillar protein composition and, in the first place, by its myosin isoform composition. Myosin is the motor of the muscle contraction—that is, it is able to convert the chemical energy of ATP into mechanical energy (work). The myosin molecule, a hexameric protein, comprises two heavy chains (MyHC) and four light chains (MyLC): both MyHC and MyLC exist in several isoforms. Four MyHC isoforms are expressed in the trunk and limb skeletal muscles of adult mammals: slow, 2A, 2X, and 2B. MyHC-2B, however, is not expressed in human muscle.2 In addition, other isoforms are expressed in specialised muscles—for example, extraocular MyHC and masticatory MyHC are expressed in extrinsic eye muscles and jaw muscles, respectively. During development respiratory muscle fibres, like all other skeletal muscle fibres, express developmental isoforms: embryonic MyHC and neonatal MyHC.1
MyHC isoforms are generally used for fibre type classification because:
-
they are the main determinants of the contractile properties;
-
MyHC is the most abundant protein; and
-
several methods used to identify fibre types, including histochemical mATPase reaction to monoclonal antibody and polyacrylamide gel electrophoresis, are based on MyHC isoforms.
Taking into account the MyHC isoform composition, muscle fibres are classified as slow (containing MyHC slow) or fast 2A, fast 2X and fast 2B (containing MyHC 2A, 2X or 2B, respectively; table 1). In human muscle fibres the maximum shortening velocity (fig 1A) and the rate of ATP hydrolysis (fig 1B) progressively increase from slow to 2A and 2X fibres.1 The tension developed during isometric contractions is generally lower in slow than in fast fibres, although this difference is much lower than for shortening velocity.4
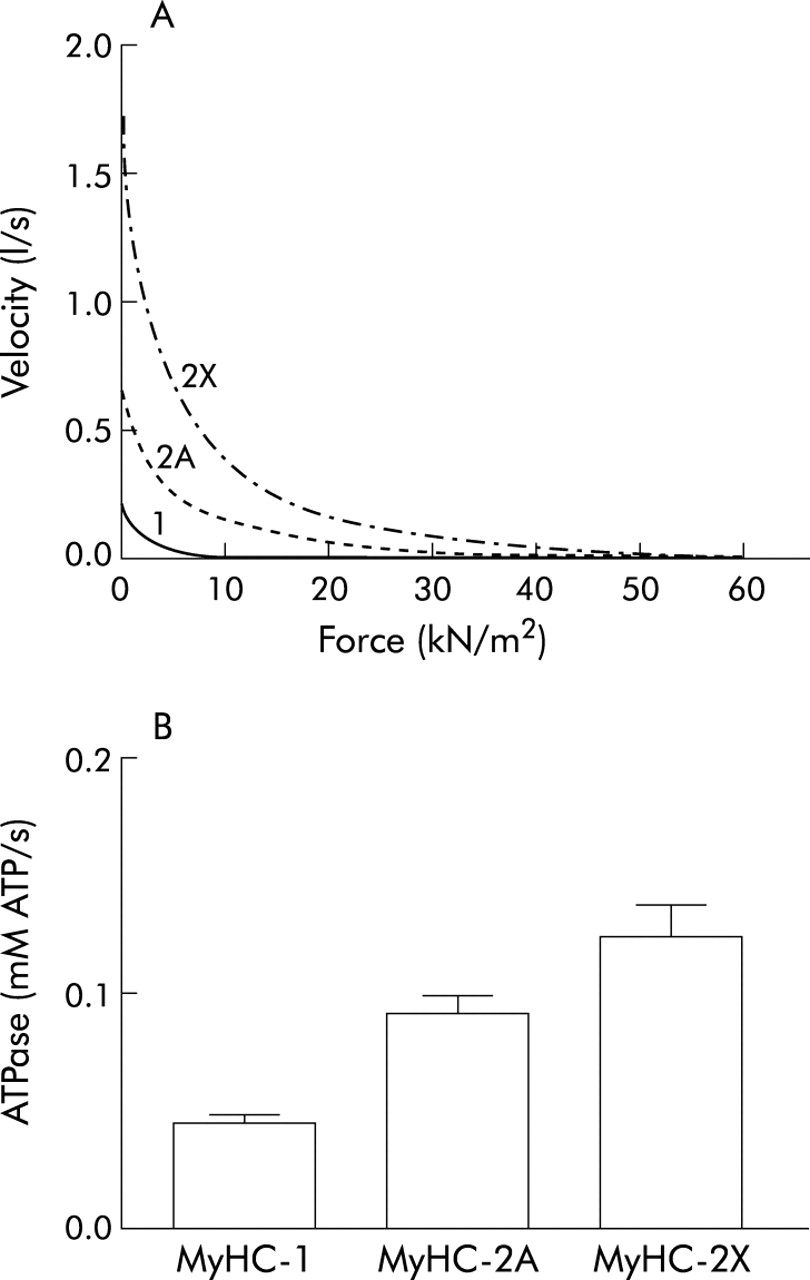
Contractile and energetic properties of single muscle fibres from human muscles (vastus lateralis). (A) Force-velocity relationships of three representative single muscle fibres containing MyHC-1 (continuous line), MyHC-2A (dotted line) and MyHC-2X (dashed line) isoforms. Velocity and force increase from slow→2A→2X. (B) Mean values of the rate of ATP consumption during isometric contractions (ATPase activity) of three single fibres containing MyHC-1, MyHC-2A and MyHC-2X isoforms. The ATPase activity of the fibre types also increases from slow→2A→2X.
Fast and slow fibres also differ in the time course of the contractile response—that is, during contraction the increased tension reaches a peak and declines faster in fast than in slow fibres (table 1). The time parameters of the contraction are determined by (1) the rate at which calcium is released and taken up from the sarcoplasmic reticulum, (2) the sensitivity of the myofibrils to calcium, and (3) the rate at which myosin forms force-generating bonds (cross bridges) with actin. In turn, the rate of calcium release and uptake depends on the properties of the sarcoplasmic reticulum: the density of release units and the density of the calcium pumps are higher in fast fibres than in slow fibres5–8 and specific isoforms of the calcium pump (SERCA) are expressed in slow and in fast fibres.9
Slow fibres are generally more resistant to fatigue than fast fibres—that is, they can maintain their contractile performance for a longer time (table 1). The resistance to fatigue depends on the balance between energy production and utilisation. The consumption of ATP for generating mechanical power output and also for control of ion (initially the calcium concentration) is matched by ATP production. ATP production follows two main metabolic routes: (1) anaerobic glycolysis which generates one ATP and two lactate or pyruvate molecules from one glucose molecule and (2) aerobic oxidation in the mitochondria where 18 ATP molecules are produced from one acetate residue (which derives either from fatty acid β oxidation or from pyruvate decarboxylation) with only carbon dioxide and water as by-products. Aerobic oxidation requires oxygen so there is a well developed capillary network around the muscle fibres and myoglobin to store oxygen in the fibres. Slow fibres are resistant to fatigue because they consume little ATP and this consumption can be completely matched by aerobic oxidative production of ATP; thus, cytosol composition is little altered by contractile activity. By contrast, the anaerobic metabolic power is very high in fast fibres; however, cytosol acidification due to by-products of anaerobic glycolysis (lactate molecules) inhibits the contractile response and causes fatigue. Consumption of ATP for contraction of fast fibres is so high that it cannot be matched by mitochondrial ATP production and a large increase in the inorganic phosphate concentration follows contractile activity. Accumulation of lactate and inorganic phosphate induces fatigue, so the loss of contractile performance is quicker in fast fibres than in slow fibres.
FIBRE TYPES IN RESPIRATORY MUSCLES
The presence of slow and fast fibres in respiratory muscles reflects their functional tasks. While quiet breathing uses mainly slow fibres, fast muscle fibres are recruited specifically when the breathing rate increases—a shift in recruitment from slow to fast muscle fibres during tachypnoea has been demonstrated in the diaphragm and parasternal muscles of the rabbit.10,11 The proportion of fast fibres in respiratory muscles is higher in small laboratory animals (mice and rats) than in humans or in large mammals: this may reflect the ventilation rate which decreases from small to large animals.12,13 Based on their pattern of activity the respiratory muscles can be classified into two main groups. During inspiration electromyographic (EMG) activity can be recorded in the diaphragm, scalenus, sternomastoid, parasternal, internal intercostal, and external intercostal muscles while during expiration the internal intercostal and abdominal muscles are active. During quiet breathing most of the respiratory work is carried out by the diaphragm and other respiratory muscles (accessory respiratory muscles) become important only when required by increased ventilation depth. The diaphragm is therefore engaged in a continuous rhythmic activity which does not permit any pause to rest, so the diaphragm fibres must be very resistant to fatigue. Besides ventilation, respiratory muscles are involved in other motor activities including coughing, talking and singing; these activities are phasic and occasional. Some respiratory muscles also have a postural role which implies more sustained activity.
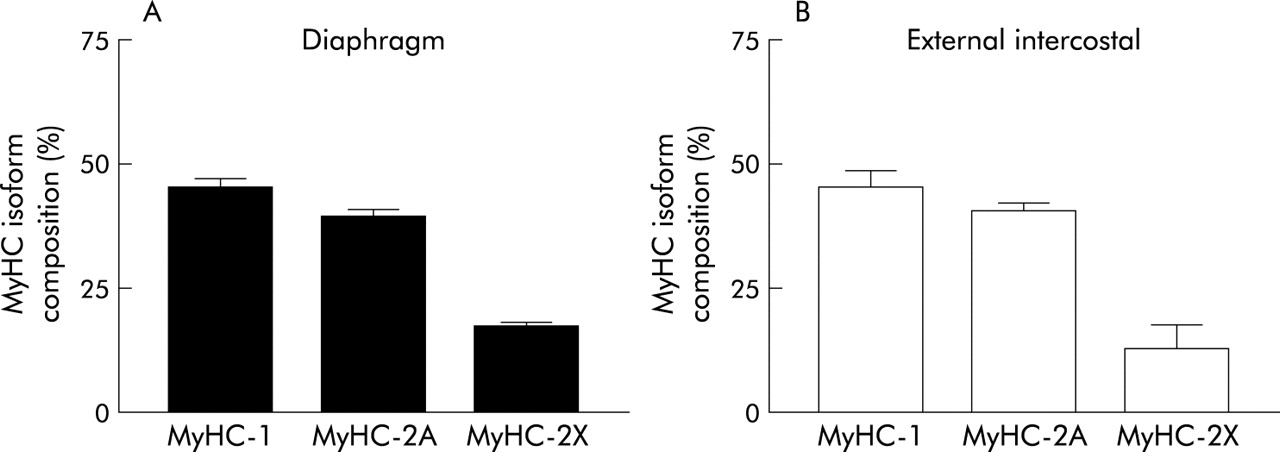
The best estimates of fibre type distribution in the adult human diaphragm indicate about 55% slow fibres, 21% fast oxidative, and 24% fast glycolytic.14–16 The distribution of myosin isoforms shows that slow myosin accounts for less than 50% of the total while myosin 2A accounts for about 40% (fig 2A). The discrepancy in fibre type distribution might be explained by the different size of each fibre type.

MyHC isoform composition of human respiratory muscles in (A) the diaphragm and (B) external intercostal muscle. In both muscles slow (MyHC-1) and fast (MyHC-2A + MyHC-2X) MyHCs are expressed in almost equal proportions. Data compiled from Levine et al168 for the diaphragm and D’Antona (unpublished observation) for the external intercostal muscle.
Information on intercostal muscles is more fragmentary. In humans the proportion of slow fibres is above 60% (that is, slightly higher than in the diaphragm16–20) in both the internal and the external intercostal muscles. The distribution of the myosin isoforms in the external intercostal muscles (fig 2B) is similar to that of the diaphragm, the discrepancy being due to the larger size of the fibres containing fast myosin.
When compared in a given animal species, fast 2X and 2B fibres have a lower resistance to fatigue and lower aerobic oxidative power than slow and fast 2A fibres (table 1), as indicated by the lower activities of succinate dehydrogenase or citrate synthase, both of which are key enzymes in mitochondrial metabolism.21–23 This difference is also present in respiratory muscles (rat diaphragm,24 cat diaphragm25), although there is a large variability in each fibre type, particularly 2X fibres. Significant differences are seen in the activity of aerobic oxidative enzymes in different species. Oxidative metabolism, measured by succinate dehydrogenase activity, is higher in smaller animals in direct proportion to the respiratory rate and the fraction of fast fibres12,26 and inversely proportional to the expression of slow myosin.13 Thus, the respiratory muscles of small animals are not only faster but also have more aerobic oxidative activity than those of large animals. The increased level of aerobic oxidative metabolism in respiratory muscles results in the increased resistance to fatigue which is required by the continuous rhythmic activity. Fatigue in respiratory muscles limits physical performance as shown in athletes and in patients with chronic obstructive pulmonary disease (COPD).27,28
Diaphragm fibres generally have a smaller cross sectional area than limb muscles. As the number of capillary vessels surrounding each fibre is similar, the diffusion distance is reduced which makes the oxygen supply more efficient in the diaphragm than in other muscles.16 An inverse relation has been reported between aerobic oxidative enzyme activity and cross sectional area.29 This might improve oxygen diffusion and contribute to the increased resistance of the diaphragm to fatigue.
Recent studies have also revealed specialised features of excitation contraction coupling in diaphragm fibres compared with other respiratory muscles and hind limb muscles. The response to agents such as caffeine, which induce calcium release without membrane depolarisation, is definitely greater in the diaphragm than in other fast muscles and this responsiveness finds its molecular basis in the expression of two isoforms of the sarcoplasmic reticulum calcium release channels—RyR1 and RyR3.30,31 Whereas RyR1 (which is present in all skeletal muscles) mediates calcium release induced by membrane depolarisation, RyR3 (which is expressed only in the diaphragm) mediates calcium induced calcium release and is more responsive than RyR1 to caffeine stimulation of calcium release.30–32 The advantage given to diaphragm fibres by this specialised arrangement is still not clear, although the greater responsiveness of the diaphragm to compounds such as methylxanthine which stimulate muscle contractility can be explained by the presence of RyR3.
DEVELOPMENT
Perinatal development in humans is characterised by a significant increase in diaphragm contractile performance, as shown by the positive correlation between both transdiaphragmatic pressure (Pdi) and maximal inspiratory pressure (Pimax) and the post-conceptional age over a range of 32–44 weeks.33,34 A similar developmental increase of the contractile force has been measured in vitro on strips of diaphragm from rat35 and hamster.36 Because of the delayed development of small rodents compared with humans, the increase in diaphragm strength can be easily observed in the early postnatal period. A recent study37 has provided a comprehensive picture of the mechanism behind the developmental increase in contractile force in rat diaphragm. Three distinct factors interplay—namely: (1) the increased density of myosin per sarcomere, (2) the replacement of slow and neonatal myosin with fast myosin, and (3) the higher force developed by individual myosin molecules in fibres expressing fast myosin. It is likely that similar mechanisms are relevant also for the human diaphragm where the replacement of neonatal myosin with adult slow and fast myosin occurs rapidly in the perinatal period—for example, the proportion of slow fibres increases from 9% at 27 weeks (gestational age) to 25% at term and reaches the adult level during the second postnatal year.38 The increase in slow myosin expression is accompanied by an increase in oxidative capacity evaluated by NADH dehydrogenase activity, producing enhanced resistance of the diaphragm to fatigue.38
TRAINING AND DISUSE
Contractile activity is one of the most effective stimuli which modify muscle fibre size and type.39 In humans, disuse causes a decrease in muscle mass40 which becomes evident in just a couple of weeks.41 Although work carried out in rat and mouse muscle has shown that disuse causes a shift towards a faster muscle phenotype,42 a clear transformation from slow to fast fibre type has only been observed in humans after strict immobilisation43 while in other models of disuse (such as space flight and bed rest) the shift is hardly significant.44–46 Interestingly, muscles of paraplegic subjects develop a uniform fast fibre composition within 6–12 months of injury.47
The two main types of muscle exercise—endurance training (long sessions of submaximal contractions) and resistance training (short sessions of maximal contractions48)—are generally believed to result in different skeletal muscle adaptations. Endurance training does not increase muscle mass and force49 and enhances the activity of aerobic enzymes,50 whereas resistance training causes a significant increase in muscle mass and strength51 with an uncertain effect on mitochondrial metabolic enzymes.52,53 In human limb muscles both endurance and resistance training result in transformation of fibres from type 2X to 2A,54–58 whereas the opposite transformation (from 2A to 2X) has not been reported in any exercise paradigm58,59 and would occur only in disuse and immobilisation. Transformation in the direction from type 1 to 2A has been shown following intense sprint training54,56,57 and transformation from 2A to 1 has been reported following endurance training.60
Respiratory muscle fibres do not undergo changes due to training and inactivity in the same way as limb muscle fibres. It has been questioned whether respiratory muscles can be trained, and the chronic activity of respiratory muscles seems to suggest that no real disuse can occur. The observation that ventilation during strenuous exercise never reaches the level of maximal voluntary ventilation and that ventilation can be increased even when subjects are exercising above the anaerobic threshold has, for a long time, led to the conclusion that the respiratory system is “built for exercise”61 and that respiratory muscles cannot be trained. However, interest in the response of respiratory muscles to exercise has grown significantly since it was shown that respiratory muscle fatigue can occur during heavy endurance exercise62 and that impairment in the performance of respiratory and limb muscles in patients with COPD is a relevant factor in determining the reduction in exercise capacity and quality of life.63
In the rat the aerobic capacity of respiratory muscles can increase following endurance training of the whole animal,64–68 notwithstanding earlier evidence showing an opposite effect.69–71 The increase in aerobic capacity was observed not only for the costal diaphragm but also for the crural diaphragm,66 parasternal,66 and external intercostal70 muscles. Interestingly, the increase occurs in all fibre types (1, 2A and 2X64,65,72 and is not caused by a shift in the composition of the fibre type. Hardly any change in fibre type composition following endurance training was, in fact, observed.73,74 This is still further evidence that, in respiratory muscles, metabolic enzymes and myofibrillar proteins are not strictly coupled as shown in limb muscles (see above). It has been pointed out, however, that the response of aerobic metabolism of respiratory muscles to endurance training,72 although clear, is less marked than in limb muscles.75 Fibre cross sectional area decreases following endurance training, and this may support the observed increase in aerobic enzyme activity64,65 determining an increase in the concentration of mitochondria.
In humans inspiratory muscle training has become an important part of the pulmonary rehabilitation programme for COPD patients with inspiratory muscle weakness.76 The effect of inspiratory muscle training on exercise performance has yet to be established,76 and there is still much debate as to whether endurance exercise performance is enhanced by respiratory training in athletes77,78 and in subjects with COPD.76,79
An increase in diaphragm shortening velocity can be induced by a moderate training programme but it is unlikely that this change reflects structural or biochemical adaptation of the muscle fibres.80 There is little direct information available on structural changes in human respiratory muscle fibres in relation to respiratory training. A recent study has shown that the functional improvement caused by a 5 week inspiratory training programme in a group of patients with COPD was associated with increases in the proportion of slow fibres (by approximately 38%) and in the size of the fast fibres (by approximately 21%) in the external intercostal muscles.81
Notwithstanding the fact that respiratory muscles are chronically active, adaptation to a reduced mechanical load can actually occur and can be responsible for the problems encountered in patients being weaned from mechanical ventilation. Animal studies have shown that a decreased diaphragm force is already observed after 12 hours of mechanical ventilation and diaphragmatic myofibril damage after 3 days. In diaphragm fibres insulin-like growth factor 1 (IGF-1) and MyoD/myogenin messengers are downregulated and oxidative stress and protease activity are increased; fibre atrophy appears after 1–2 days of controlled mechanical ventilation.82
AGEING
The force, mass, and structure of skeletal muscle are affected by ageing. In humans the force of skeletal muscle in vivo significantly decreases (20–40%) with ageing,83–89 the most likely explanation of such decrease being the loss of muscle mass. Muscles can lose 20–30% of their mass by the age of 70.87,90 The decrease in muscle mass is believed to depend on a decrease in both the size and the number of muscle fibres. The size of skeletal muscle fibres is reduced in subjects over 70 years with a selective atrophy of fast fibres.85,91 Partial denervation of muscle fibres occurs with ageing92,93 and, notwithstanding partial re-innervation, both motor units92 and muscle fibres90,93,94 are lost in ageing. Slow motor units re-innervate muscle fibres more efficiently than fast motor units and therefore considerably increase in size.92,93 This would suggest a general shift towards the slow fibre phenotype. There are, however, several studies showing either no change in fibre type distribution95,96 or even an increase in fast fibres.97–99 A possible explanation for this is represented by the concomitant muscle disuse related to the general reduction in motor activity typical of ageing. The increased proportion of slow fibres does not make muscle metabolism more oxidative. Maximal oxidative power decreases with ageing in relation to the deterioration in mitochondrial function.100,101
There is limited information available on the effect of ageing on respiratory muscles, particularly in humans. Moreover, the fact that respiratory muscles must be chronically active suggests that ageing might differentially affect respiratory and limb muscles and it is therefore difficult to apply information on limb muscles to respiratory muscles. That ageing does affect respiratory muscles is supported by the significant reduction in diaphragm strength observed both in small mammals102,103 and in humans.104,105 Consistent with the uncertain effect of ageing on limb muscles, van Lunteren et al106 found a small but significant shift towards type 2B fibres in the diaphragm of old rats while other workers103,107 have reported no change. The lack of single muscle fibre atrophy observed in limb muscles finds its counterpart in the diaphragm with no change in the cross sectional area of single fibres in old rats.106,107 Finally, the uncertain effect of ageing on metabolic enzymes is consistent with the lack of significant changes in oxidative capacity of the diaphragm observed in old rats.107–109 No specific ageing effect on respiratory muscles has therefore been found to date, in spite of their peculiar functional role.
MUSCULAR DYSTROPHIES
The diseases which primarily affect the respiratory muscles are muscular dystrophies. Mutations of genes coding for cytoskeleton, sarcolemma, the nuclear envelope, and extracellular matrix cause early death of muscle fibres which is probably mediated by a loss of membrane integrity. For unknown reasons the muscle groups which mark the onset of the disease vary among the muscular dystrophies but are mostly represented by limb muscles. Respiratory insufficiency occurs late in patients with Duchenne, Becker and limb girdle dystrophies, being one of the most common causes of death,110 and is very infrequent in patients with facio-scapulo-humeral muscular dystrophy.111 Congenital myopathies such as Ulrich’s congenital myopathy112 which is characterised by early severe respiratory impairment is an exception. The specialisation and peculiar pattern of activity of respiratory muscle fibres might account for their variable and either late or lack of involvement in muscular dystrophies.
There is no direct information available on changes in skeletal muscle fibres from respiratory muscles in humans with muscular dystrophy. Indirect evidence of pseudo-hyptrophy in the diaphragm of patients with Duchenne’s muscular dystrophy has been obtained in a recent study which combined ultrasonographic measurement of diaphragm thickness with recording of transdiaphragmatic pressure.113 At variance with the human counterpart, the mice model of Duchenne’s muscular dystrophy (mdx mice) is characterised by primary involvement of the diaphragm114 while the limb muscles do not have severe functional deficits comparable to the clinical symptoms observed in boys with the condition.115 The diaphragm of mdx mice contains regenerating fibres which express developmental MyHC and has a general shift towards slower fibre types (1 and 2A) at the expense of the fastest fibre types (2X and 2B) and a very significant loss of specific force.114 Impaired diaphragmatic function has also been seen in two models of limb girdle muscular dystrophy: Bio 14.6 hamsters lacking δ-sarcoglycan116 and α-sarcoglycan null mice.117 In spite of the different involvement of respiratory and limb muscles, murine models of human muscular dystrophies provide useful information on their pathogenesis and have been extensively used to test possible treatments.
Attempts to rescue muscles affected by muscular dystrophy are currently in progress along three main directions:118
-
Gene therapy which aims to deliver the mutated gene to dystrophic muscle fibres by adenovirus and adeno-associated viruses. Intramuscular injection of such vectors has determined expression of mini-dystrophin (the full length dystrophin is too large to be successfully carried) in mdx mice119,120 and restored the expression of α-sarcoglycan121 in α-sarcoglycan null mice and of δ-sarcoglycan122 in Bio 1.6 hamsters. However, the lasting transgene delivery to the whole body without immune reaction remains a major problem.123
-
Pharmacological treatment based on the observation that most dystrophin mutations occur in a region of the molecule that is not strictly necessary for function but that just stops transcription before a viable molecule can be synthesised. Gentamicin124 and oligonucleotides125 have been successfully used to determine exon skipping and to enable transcription of a dystrophin molecule of almost normal length and sequence. The effectiveness of gentamicin treatment has recently been challenged126 and clinical trials have so far had negative results.118
-
Cell therapy which aims to deliver to dystrophic muscles satellite cells or pluripotent stem cells that can differentiate into skeletal muscle, either to repair degenerating skeletal muscle cells or to generate new skeletal muscle cells. The limited self-renewal and migratory capacity of satellite cells and myoblasts have prevented significant success so far. The intra-arterial delivery of a new class of vessel-associated fetal stem cells (the mesoangioblasts) in α-sarcoglycan null mice has recently shown a dramatic amelioration of the morphology and function of the downstream muscles,127 opening new prospects for cell therapy.
Respiratory muscles have not so far been treated in animal models of muscular dystrophies. In small animals such as mice the respiratory muscles are difficult to reach either by intramuscular injection or intra-arterial delivery. However, in humans, delivery is not a problem and respiratory muscles will be treated in addition to limb muscles when human treatments have been established.
MALNUTRITION
Prolonged malnutrition/undernutrition causes a loss in body weight in parallel with skeletal muscle wasting and the respiratory muscles are not spared, the diaphragm often being the most affected of the respiratory muscles. The diaphragm weight loss—which, at least in rodents, appears to be related to sex being more marked in males128—is proportional to that of the other skeletal muscles (the diaphragm atrophies as much as the limb muscles129) or even exceeds the loss of body weight as seen from necroscopic studies of undernourished subjects.130 It is uncertain whether the loss in muscle mass is due to a reduction in fibre cross sectional area or to a decrease in fibre number.131,132 Some studies have found the fibre number to be reduced by starvation,133 others have reported no changes131,132 and others, while not excluding a possible effect of food deprivation on total cellularity, have shown a primary effect of undernutrition on fibre size.129
Several studies have shown a selective effect of undernutrition on fast fibres in both respiratory134–136 and hind limb muscles131,137,138 which appear to atrophy exclusively134,135 or to greater extent129,131,136,139 than slow fibres. The change in the composition of diaphragmatic fibres towards a slower phenotype induced by undernutrition134 might be partly attributable to the transition from fast to slow fibres caused by the increase in circulating glucocorticoids resulting from starvation140 and partly to the fact that slow fibres are more active than fast fibres.141
The loss of muscle mass and the greater effect of undernutrition on fast fibres have a considerable effect on the contractile and fatigue properties of the diaphragm. A large reduction in both peak twitch force (Pt) and peak tetanic tension (Po) of diaphragm strips without changes in specific force (Pt and Po corrected for diaphragm muscle weight or cross sectional area) as well as a prolongation of twitch half relaxation time (RT0.5, time for Pt to fall to 0.5 of maximum) and an increase in Pt/Po have been reported.129,142–145 The absence of starvation induced changes in specific force suggest that the fall in force generation observed in the diaphragm should be attributed exclusively to the reduction in muscle mass and not to coupled alterations of the myofibrillar machinery. Furthermore, the observed changes in RT0.5 and Pt/Po might relate to the increased proportion of slow fibres.129,144,145 Whereas resistance to low frequency fatigue increases,129,144–146 resistance to high frequency fatigue is reduced144 and these changes can be detected by assessing diaphragm contractility via indirect phrenic stimulation.142,143
GLUCOCORTICOIDS AND BETA AGONISTS
Several pharmacological compounds can modify the size and/or type of skeletal muscle fibres, of which glucocorticoids and β agonists are relevant to the treatment of respiratory disease.
The continuous use of large doses of glucocorticoids causes muscle atrophy and weakness, a condition known as steroid myopathy which is well characterised in humans and in animals. The contractile impairment can be sufficiently severe to reduce motor activity and ventilatory performance.147,148 Steroid myopathy is characterised in both animal and human muscles by fibre atrophy which predominantly affects fast fibres, by a shift from fast to slow myosin isoform expression, and by increased variation in fibre cross sectional area with several angular atrophic fibres and necrotic fibres.148,149 Since a possible explanation for the relative resistance of slow fibres is their more sustained activity, one might anticipate that muscle fibres are less affected in respiratory muscles than in limb muscles. The typical signs of steroid myopathy are, however, also well documented in the diaphragm of rats149,150 and hamsters151 and in the intercostal muscles of rats.149
The chronic administration of β agonists to relax airway smooth muscle causes hypertrophy of skeletal muscles which is more pronounced in fast than in slow fibres, and a transition from slow to fast myosin isoforms. The effect is seen in limb muscles and respiratory muscles of various animal species.152–154 Beta agonists therefore improve the contractile performance of respiratory muscles as shown in hamsters155 and rats.156 It is worth remembering that enhanced contractility of the diaphragm also follows the acute administration of some β agonists such as salbutamol.156
Interestingly, the acute effect of β adrenergic activation has opposite effects in fast and slow muscles,157 the amplitude and duration of the twitch being increased in fast muscles and decreased in slow muscles. This implies that both chronic and acute administration of β agonists have variable effects on slow and fast fibres. The distribution of β2 adrenoreceptors is heterogeneous. In hind limb muscles there are 85% β2 receptors and 15% β1 receptors while, in the diaphragm, only β2 receptors are present. The diaphragm would therefore be expected to be more sensitive to β2 agonists than hind limb muscles. Furthermore, the density of β2 receptors is greater in slow fibres than in fast fibres.158
CHRONIC OBSTRUCTIVE PULMONARY DISEASE
One of the first complaints of patients with COPD is reduced exercise tolerance, which is caused partly by the structural and functional impairment of skeletal muscles. Analysis of biopsy specimens from the quadriceps femori of patients with COPD has shown significant fibre atrophy with a reduction in the proportion of slow fibres accompanied by an increase in fast 2X fibres.159,160 In contrast, no changes occur in the biceps brachii of COPD patients compared with age matched controls,161 and handgrip strength is less reduced than quadriceps strength.162 A significantly increased expression of MyHC-2X and of fast MyLC isoforms have been observed in the vastus lateralis of COPD patients compared with controls.163 Interestingly, the capillary density is dramatically reduced and myoglobin concentration is decreased in the muscles of patients with COPD,164 and oxidative aerobic enzyme activity is also reduced.165,166 Lipofuscin is accumulated in fibres, which suggests that oxidative damage of muscle tissue may be involved in skeletal muscle dysfunction and wasting in COPD.167
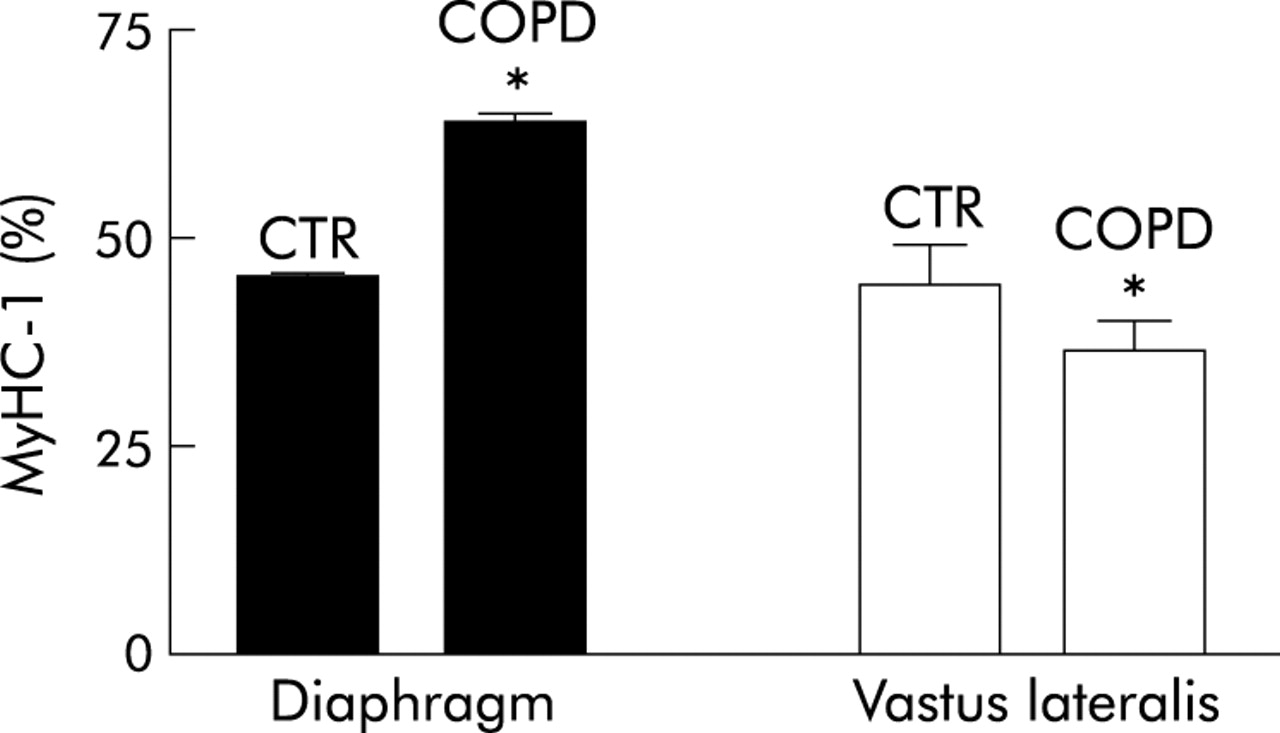
Patients with COPD have a higher percentage of slow MyHC (fig 3) and lower percentages of fast MyHC-2A and MyHC-2X in the diaphragm than controls.168 In addition, the diaphragm of patients with COPD contains a higher percentage of the slow isoforms of MyLC, troponins, and tropomyosin than the diaphragm of control subjects.168 Mitochondrial oxidative capacity relative to ATP demand (SDH/mATPase) is higher in each of the fibre types in the diaphragm of COPD patients than controls, pointing to an increased resistance to fatigue.169 Apparently, the increased functional demand induces an adaptation in the diaphragm fibres of patients with COPD. Surprisingly, developmental (embryonic and neonatal) MyHC isoforms have been found in the diaphragms of patients with severe COPD, but the significance of this remains to be elucidated.170 Ultrastructural studies have also revealed subcellular changes in the diaphragm of patients with COPD including a reduction in the mean length of the sarcomere at rest and an increase in the concentration of mitochondria.171 The changes in expiratory muscles might differ from those in the diaphragm: a decrease in force and endurance has been shown in a recent study.172

{kind=link}
{kind=link}
{kind=link}
Relative proportion of MyHC-1 isoform in biopsy samples from the diaphragm and vastus lateralis muscles of healthy subjects (CTR) and patients with chronic obstructive pulmonary disease (COPD). Data from Levine et al168 for the diaphragm muscle and Satta et al163 for the vastus lateralis muscle; *p<0.05.
HYPOXIA
Normobaric hypoxia is a common complication of several respiratory diseases. Whereas acute severe hypoxia causes respiratory failure associated with severe damage to respiratory muscle fibres within a few hours,173 chronic hypoxia induces moderate muscle fibre adaptation.
In rat limb muscles adaptation to chronic hypoxia causes a decrease in slow fibres and an increase in fast fibres and a shift towards a more glycolytic metabolism with a time course of many weeks.174–176 These changes might be attributed to inactivity and anorexia which are also induced by lack of oxygen, but they have also been seen in rats exposed to endurance training in an hypoxic environment.174 The diaphragm does not seem to differ from limb muscles in its adaptation to hypoxia, although a recent study showed that intermittent hypoxia induces adaptive responses in the diaphragm which are able to increase its tolerance to acute anoxia.177
In patients with chronic hypoxia due to COPD the proportion of fast fibres is increased and that of slow fibres is decreased,160 but it is difficult to attribute this effect with certainty to hypoxia alone. In mountaineers exposed for several weeks to altitude hypoxia, a myopathy characterised by a significant reduction in muscle fibre size (−20%) and a loss of muscle oxidative capacity (−25%) have been observed; the mitochondrial volume is reduced and lipofuscin accumulates in the fibre cytosol.178,179 These data were derived from biopsy specimens of the vastus lateralis muscles and it is not known whether comparable alterations also occur in respiratory muscles.
CONCLUSIONS
The picture of the structural and functional features of respiratory muscle fibres which emerges from the information currently available is far from complete (table 2), although indications of their specialised performance and of their adaptation to several conditions which change the functional requirements have been described in the literature. Some descriptive information is still lacking, such as that concerning age related changes or the effects of training on human respiratory muscles. The new tools of proteomics and trascriptomics have not yet been applied to the study of respiratory muscle fibres, and analysis of the signals regulating gene transcription and post-translation protein modification has yet to be undertaken.
Summary of changes occurring in respiratory muscle fibres in relation to changes in physiological conditions, diseases, and pharmacological treatments
REFERENCES
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵