Article Text

Abstract
Rationale The pathogenesis of respiratory syncytial virus (RSV) bronchiolitis in infants remains poorly understood. Mouse models implicate pulmonary T cells in the development of RSV disease. T cell responses are initiated by dendritic cells (DCs), which accumulate in lungs of RSV-infected mice. In infants with RSV bronchiolitis, previous reports have shown that DCs are mobilised to the nasal mucosa, but data on lower airway DC responses are lacking.
Objective To determine the presence and phenotype of DCs and associated immune cells in bronchoalveolar lavage (BAL) and peripheral blood samples from infants with RSV bronchiolitis.
Methods Infants intubated and ventilated due to severe RSV bronchiolitis or for planned surgery (controls with healthy lungs) underwent non-bronchoscopic BAL. Immune cells in BAL and blood samples were characterised by flow cytometry and cytokines measured by Human V-Plex Pro-inflammatory Panel 1 MSD kit.
Measurements and main results In RSV cases, BAL conventional DCs (cDCs), NK T cells, NK cells and pro-inflammatory cytokines accumulated, plasmacytoid DCs (pDCs) and T cells were present, and blood cDCs increased activation marker expression. When stratifying RSV cases by risk group, preterm and older (≥4 months) infants had fewer BAL pDCs than term born and younger (<4 months) infants, respectively.
Conclusions cDCs accumulate in the lower airways during RSV bronchiolitis, are activated systemically and may, through activation of T cells, NK T cells and NK cells, contribute to RSV-induced inflammation and disease. In addition, the small population of airway pDCs in preterm and older infants may reveal a distinct endotype of RSV bronchiolitis with weak antiviral pDC responses.
- Viral infection
- Respiratory Infection
- Paediatric Lung Disaese
- Innate Immunity
Statistics from Altmetric.com
Key messages
What is the key question?
Are populations of lung dendritic cells (DCs), which have been implicated in respiratory syncytial virus (RSV) pathogenesis in mouse models, present in the lower airways of infants with RSV bronchiolitis and do they differ between subsets of affected infants?
What is the bottom line?
We demonstrate the accumulation of conventional DCs in the lower airways and their systemic activation in severe RSV bronchiolitis, implicating pro-inflammatory DC responses in the pathogenesis, and report a failure of airway plasmacytoid DCs to increase in numbers in premature and older (≥4 months) infants with RSV bronchiolitis, suggesting a poor antiviral response as a pathogenetic factor.
Why read on?
This first description of DCs in the lower airways in RSV bronchiolitis raises the possibility of distinct endotypes, and recognition of such DC-based endotypes may enable targeting of future antiviral and anti-inflammatory therapy to the appropriate patients with bronchiolitis.
Introduction
Respiratory syncytial virus (RSV) is the leading cause of infant viral bronchiolitis worldwide, resulting in major morbidity, requiring hospitalisation of 2% of infants and necessitating mechanical ventilation in the most severely affected.1 ,2 There is no active vaccination against RSV infection and no effective specific treatment. To address these major unmet clinical needs, better understanding of the immunopathogenesis of severe RSV bronchiolitis in infants is essential.
Mouse models of RSV infection suggest that pulmonary T cells are critical for the development of RSV-induced inflammation and disease.3 ,4 Primary T cell responses are initiated by dendritic cells (DCs) that present peptide antigens bound to major histocompatibility complex (MHC) class II molecules together with selected co-stimulatory molecules to activate naive and effector T cells and to determine the type of their response. There are two major DC subsets; conventional DCs (cDCs) and interferon (IFN)α-producing antiviral plasmacytoid DCs (pDCs).
In RSV-infected mice, we have previously shown an increase in lung cDCs, coinciding with the onset of inflammation, which exhibit increased expression of the co-stimulatory molecule CD86 and of the integrin CD11b, and when isolated induce robust T cell proliferation, indicating a pro-inflammatory phenotype.5 In addition, we and others have also shown early increases in lung pDC numbers in RSV-infected mice.6 ,7 These pDCs are required to limit RSV replication6 and may also have a regulatory role limiting inflammation, following RSV infection.7
Prematurity and young postnatal age are known risk factors for severe RSV bronchiolitis8–10 and prematurity is thought to be a determinant of RSV immunopathogenesis based on differential airway neutrophil and cytokine responses.11–13 However, little is known regarding pulmonary cellular immune responses beyond neutrophils, and data on human DC phenotype and function in RSV infection are limited. Monocyte-derived DCs14 ,15 and peripheral blood cDCs16 from healthy adults upregulate maturation markers and produce pro-inflammatory cytokines and interleukin (IL)-10 upon RSV infection in vitro, while their capacity to induce T cell proliferation is decreased,15 ,16 in contrast to murine lung cDCs after RSV infection in vivo. In pDCs, but not cDCs, in vitro RSV infection induces strong IFNα production.17 In infants with RSV bronchiolitis, increases in both cDCs and pDCs have been demonstrated in the nasal mucosa;18 however, information on DC populations in the lower airways in affected infants is lacking.
Focusing on DCs, we assessed cellular immune responses in bronchoalveolar lavage (BAL) and peripheral blood samples from infants with severe RSV bronchiolitis. We hypothesised that, parallel to observations in mice, cDC and pDC populations are increased and activated in the lower airways of infants during RSV bronchiolitis. Furthermore, we explored differential DC response patterns in defined risk groups for severe RSV bronchiolitis.
Materials and methods
Study population
Using an observational case/control study, we collected BAL and peripheral blood samples between October 2010 and December 2012 from infants with RSV bronchiolitis admitted to paediatric intensive care units at the Royal Hospitals for Sick Children in Edinburgh and Glasgow. We included infants born from 24 weeks of gestation, aged <18 months, who required intubation and ventilation with a clinical diagnosis of viral bronchiolitis. For RSV diagnosis and exclusion criteria, see online supplement. The control group comprised of healthy infants without respiratory infection in the preceding two weeks, who were intubated and ventilated during planned surgery for conditions not affecting the lung.
supplementary data
Non-bronchoscopic bronchoalveolar lavage and cell isolation
BAL was performed according to a validated protocol19 and BAL cells were isolated using standard techniques. For details, see online supplement. In addition, 1–2 mL of venous blood were collected on EDTA on the day of BAL sampling. All samples were kept on ice and processed freshly within 4 hours.
Cell staining for flow cytometry
BAL and blood cells were stained using standard techniques and commercially available antihuman fluorochrome-conjugated antibodies. For details, see online supplement. Stained samples were acquired on a BD LSR Fortessa flow cytometer. FACs Diva software V.6.1 (BD Bioscience, Oxford, UK) was used for digital data acquisition and postacquisition analysis performed using FlowJo V.7.6.5 software (treestar.inc, Oregon, USA).
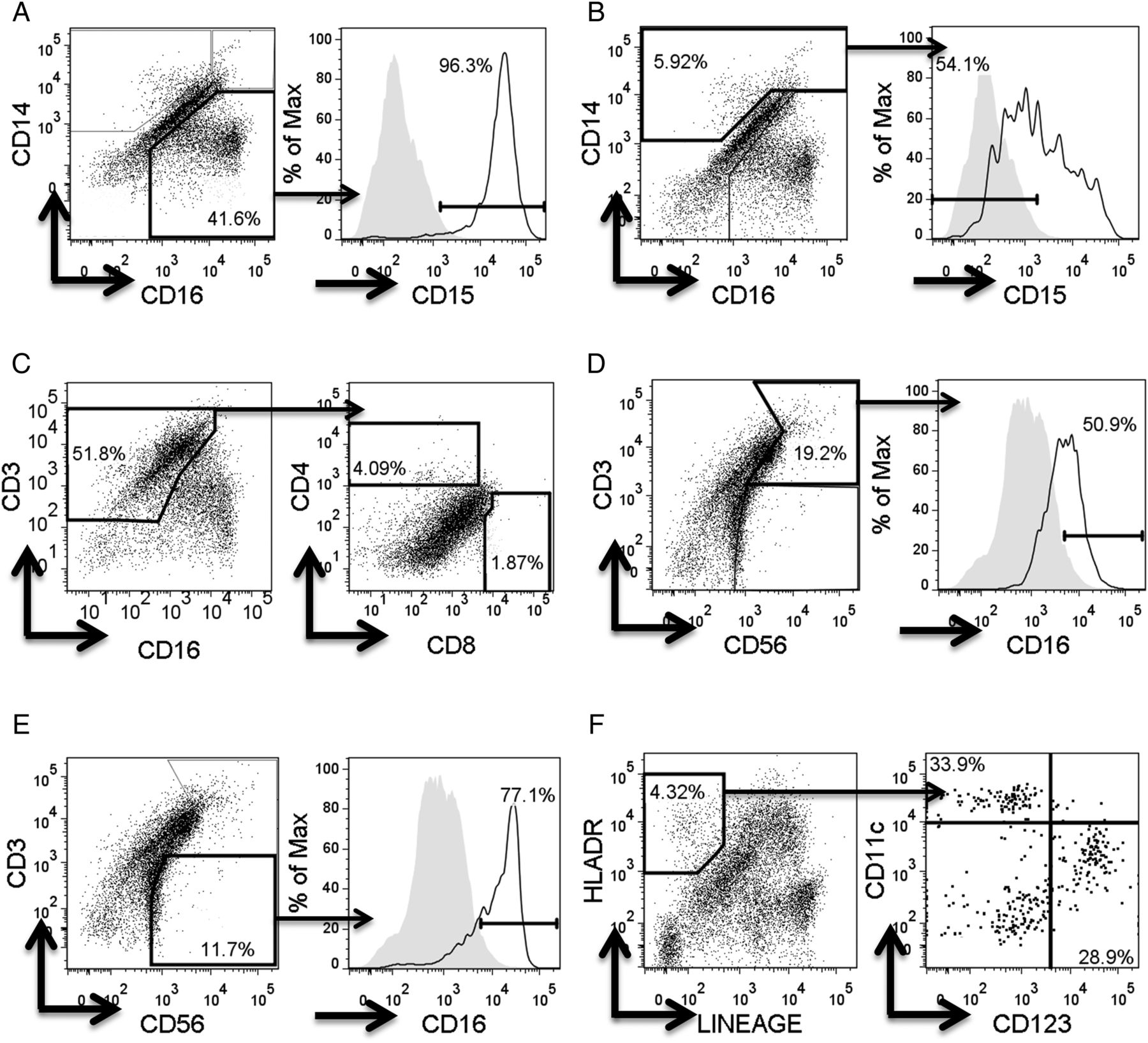
Cells were gated by forward scatter (FSC)/side scatter (SSC) to eliminate cell debris, on live cells using viability dye eFluor 780 (eBioscience), and on singlets using FSC-area versus FSC-height. All cell gates (figure 1) were defined using isotype controls.

Flow cytometry gating strategies for immune cell populations in respiratory syncytial virus bronchoalveolar lavage samples. For flow cytometry, cells were incubated with fluorochrome-conjugated antihuman antibodies and gated by forward scatter (FSC)/side scatter (SSC) to eliminate cell debris, on live cells, and on singlets using FSC area/height. All cell gates were defined using multiple isotype staining panels. The isotype staining (shaded areas) shown in the histograms is illustrative and derived from above panels. Individual immune cell populations were identified as follows; CD14negative CD16+ CD15+ neutrophils (A), CD14+ CD16+ or negative CD15negative monocyte/macrophages (B), CD3+ CD4+ helper T cells / CD3+ CD8+ cytotoxic T cells (C), CD3+ CD56+ CD16+ NK T cells (D), CD3− CD56+ CD16+ NK cells (E) and lineagenegative (containing anti-CD3, anti-CD14, anti-CD16, anti-CD19, anti-CD20 and anti-CD56), human leukocyte antigen (HKA)-DR+, CD11c+ conventional dendritic cells and CD123+ plasmacytoid dendritic cells (F).
Cytokine measurements
Cytokines (IFNγ, IL-1β, IL-2, IL-4, IL-6, IL-8, IL-10, IL-12p70, IL-13 and tumour necrosis factor (TNF)-α) were quantified using the human V-Plex Pro-inflammatory Panel 1 Kit (Mesoscale Discovery; cat. K15049D). Mean lower limits of detection were IFNγ (0.2 pg/mL), IL-1β (0.04 pg/mL), IL-2 (0.09 pg/mL), IL-4 (0.02 pg/mL), IL-6 (0.06 pg/mL), IL-8 (0.04 pg/mL), IL-10 (0.03 pg/mL), IL-12p70 (0.11 pg/mL), IL-13 (0.24 pg/mL) and TNF-α (0.04 pg/mL). Concentrations were measured in RSV bronchiolitis BAL (n=35) and serum (n=24) and in control BAL (n=8) and serum (n=6).
Statistical analysis
Data are expressed as median and IQRs. Mann-Whitney U test was used for comparisons between RSV cases and controls, preterm and term born RSV cases, and younger (<4 months) and older (≥4 months) RSV cases. The relationship between cytokines (after logarithmic transformation) and cDC/pDC numbers was examined using Spearman correlation coefficients (r). p Values <0.05 were considered statistically significant. All data were analysed with GraphPad Prism V.5.0 (GraphPad Software, San Diego, USA).
Results
Patient demographics
We recruited 37 infants mechanically ventilated for RSV bronchiolitis. No significant differences in chronological age (11.8 (IQR 4.5–18.9) vs 5.3 (IQR 3.5–10.3) weeks, p=0.13) or corrected age (42.4 (IQR 39.2–51.5) vs 44.6 (IQR 41.2–51) weeks, p=0.55) were found between the 9 preterm and 28 term born infants. The first seven cases were used to establish the staining protocol and are not included in the analyses. One BAL sample was excluded as an outlier with total BAL cell numbers >2 SD above the mean. Of the remaining 29 cases, 24 (82.8%) had sufficient BAL quality for flow cytometric analysis. In addition, 14 controls were recruited, who were intubated for surgery including laparoscopy, inguinal herniotomy, renal pyeloplasty, ileal stoma closure, hip osteotomy and cleft palate/lip repair. These controls were older than the RSV cases (31.8 (IQR 19–60.6) vs 6.7 (IQR 4–14.3) weeks, p=0.0005). Only four controls (28.6%) had BAL samples of sufficient quality and cellularity for flow cytometry. Furthermore, peripheral blood samples from 19 infants with RSV bronchiolitis and 5 controls were collected and analysed by flow cytometry.
Of the 24 infants with RSV bronchiolitis whose BAL samples were analysed by flow cytometry, 9 were born prematurely (<37 weeks, 6/9 male) and 15 at term (≥37 weeks, 8/15 male). When stratified by postnatal age, 17 infants were <4 months old (8/17 male, 6/17 preterm) and 7 infants were 4 months or older (6/7 male, 3/7 preterm).
BAL samples
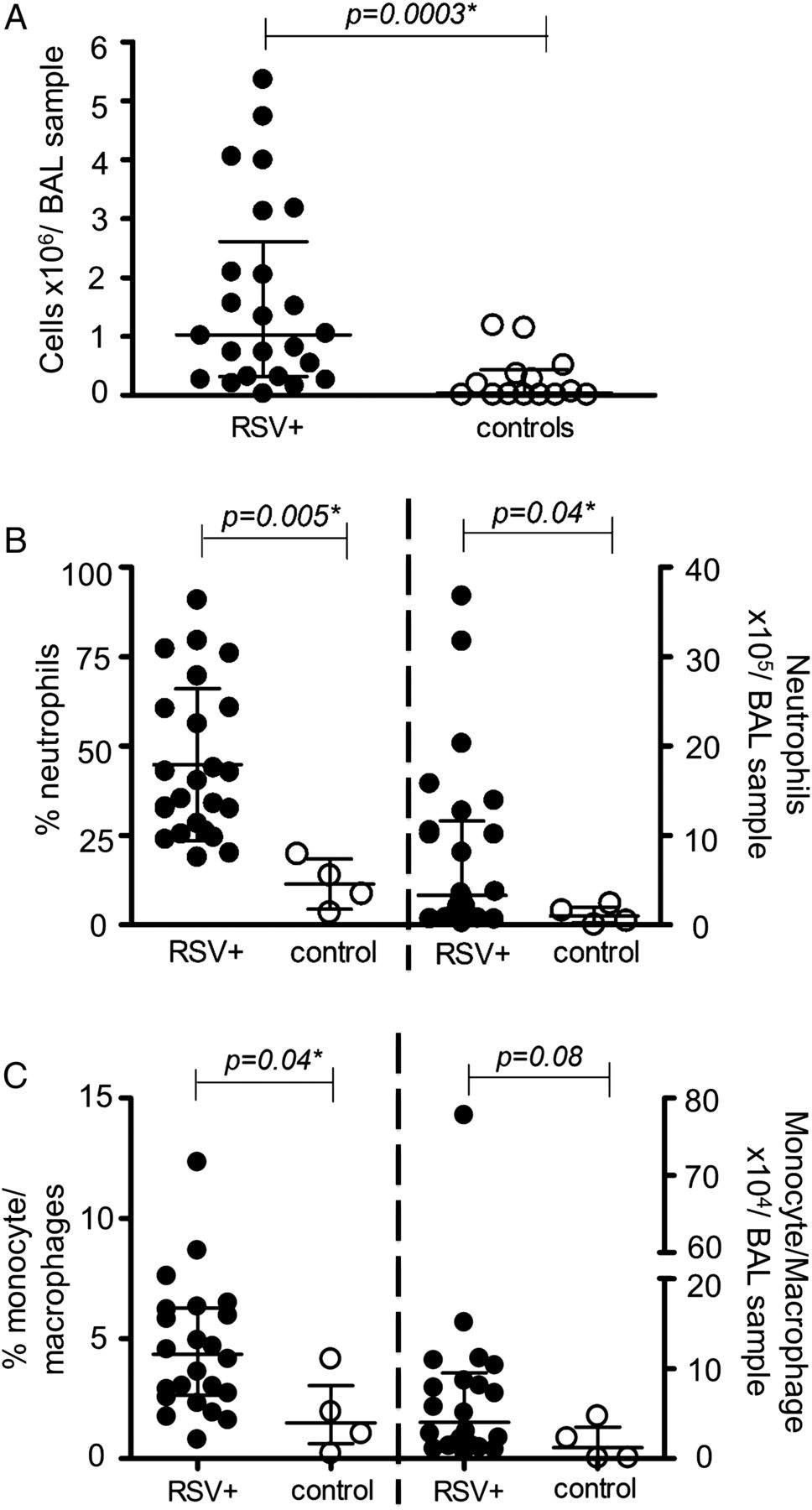
BAL volumes recovered from 29 infants with RSV bronchiolitis (2.8±1.2 mL) were similar to those from the 14 control subjects (2.2±1.2 mL). Total BAL cellularity was significantly higher in infants with RSV bronchiolitis compared with controls (figure 2A).

Total cells, neutrophils and monocyte/macrophages in respiratory syncytial virus (RSV) and control bronchoalveolar lavage (BAL) samples. Total cellularity was assessed in RSV-BAL (closed circles, n=24) and control-BAL (open circles, n=14) (A). Percentages and numbers of neutrophils (B) and of monocytes/macrophages (C) were assessed in RSV-BAL (closed circles, n=24) and control-BAL (open circles, n=4). Data are expressed as medians (IQR) of cells/BAL sample. p Values were determined by Mann-Whitney U tests.
Neutrophil and monocyte/macrophage populations in BAL and peripheral blood
Neutrophils (see figure 1A and online supplementary figure S1A) were the predominant airway leucocytes in RSV-BAL samples consistent with previous reports.11 Their percentage and numbers in the airways were significantly higher in infants with RSV bronchiolitis than in controls (figure 2B). Percentages of peripheral blood neutrophils were also significantly higher in RSV bronchiolitis cases compared with controls (37.0 (IQR 30.9–45.9)% vs 21.1 (IQR 9.44–24.3)%, p=0.005).
Monocytes and macrophages (see figure 1B and online supplementary figure S1B) were also found at a significantly higher percentage in RSV-BAL than in controls (figure 2C), but without differences in peripheral blood (additional data in online supplement).
T cells in BAL and peripheral blood
Both CD4+ and CD8+ T cells (see figure 1C and online supplementary figure S1C) were present in the airways of RSV cases and accounted for 0.5%±0.6% and 2.1%±2.3% of BAL cells, respectively. There were no significant differences between RSV bronchiolitis and controls in BAL CD4+ and CD8+ T cell percentages or numbers; however, the CD8/CD4 ratio was 10-fold higher in RSV cases than controls (3.1 (IQR 1.7–7.6)% vs 0.3 (IQR 0.2–2.9)%, respectively, p=0.08), without reaching statistical significance. In the blood, CD4+ T cell percentages were significantly lower in RSV cases compared with controls (19.5 (IQR 14.2–24.2)% vs 28.7 (IQR 23.9–29.5)%, p=0.02), while CD8+ T cell percentages increased without reaching statistical significance (8.81 (IQR 5.3–12.2)% vs 5.7 (IQR 4.7–7.6)%, p=0.08).
NKT and NK cells are increased in BAL in infants with RSV bronchiolitis
Furthermore, we assessed percentages and numbers of NK T cells (see figure 1D and online supplementary figure S1D) and NK cells (see figure 1E and online supplementary figure S1E). In RSV cases, NK T cell (figure 3A) and NK cell (figure 3B) percentages in the BAL were 5.2-fold and 8-fold higher, respectively, than in controls. Apparent increases in BAL NK T cell and NK cell numbers failed to reach statistical significance. In blood samples, NK T cells or NK cells did not differ between RSV cases and controls (additional data in online supplement).

NKT and NK cell percentages are increased in respiratory syncytial virus (RSV)-bronchiolitis compared with control bronchoalveolar lavage (BAL) samples. Percentages and numbers of NK T cells (A) and NK cells (B) were determined in RSV-BAL (closed circles, n=24) and control-BAL (open circles, n=3). Data are expressed as medians (IQR) of cells/BAL sample. p Values were determined by Mann-Whitney U tests.
DCs are increased in BAL and decreased in peripheral blood in infants with RSV bronchiolitis
DCs were characterised as lineagenegative (fluorescein isothiocyanate (FITC) channel), human leukocyet antigen (HLA)-DR+ cells, with cDCs expressing CD11c+ and pDCs expressing CD123+ (see figure 1F and supplementary figure S1F). Alveolar macrophages, which are highly autofluorescent in the FITC channel, were excluded by gating out the lineage+ cell population. We found 2.2%±1.2% of lineagenegative, HLA-DR+ cells in RSV-BAL with cDCs accounting for 1.2%±1.5% and pDCs for 0.2%±0.4% of total BAL cells.
Numbers of cDCs were significantly higher in RSV-BAL compared with controls (figure 4A), while pDC numbers did not differ significantly between RSV and control BAL (figure 4A). In contrast to elevated cDC numbers in the airways, cDC percentages were significantly lower in the peripheral blood of RSV cases compared with controls, as were pDC percentages (figure 4B).
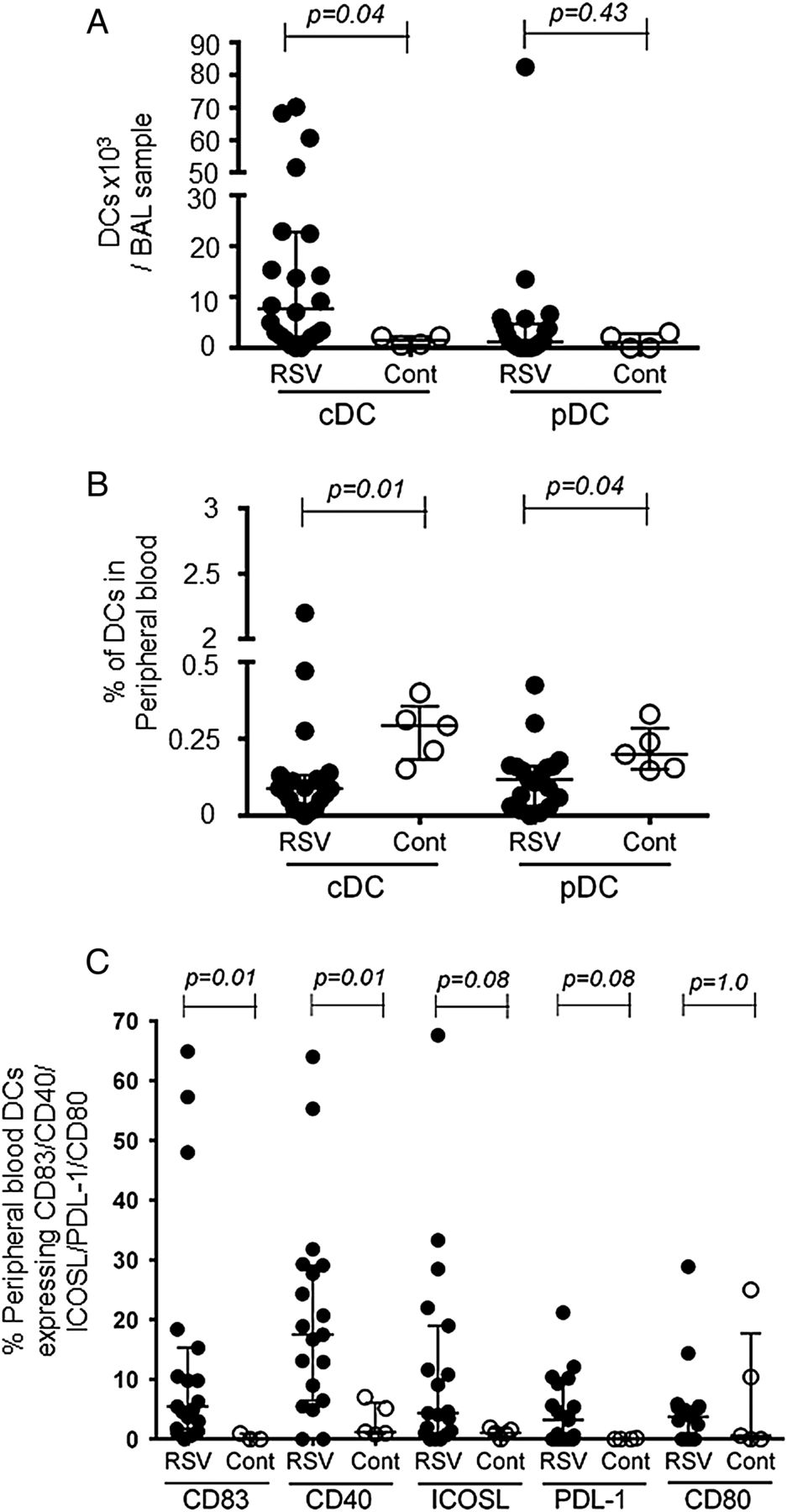
Airway conventional dendritic cell (cDC) numbers increase and percentages of peripheral blood dendritic cells (DCs) decrease in infants with respiratory syncytial virus (RSV) bronchiolitis. DCs were characterised by flow cytometry as lineagenegative, HLA-DR+ cells, with cDCs expressing CD11c+ and plasmacytoid dendritic cells (pDCs) CD123+. (A) Numbers of cDCs and pDCs in RSV-bronchoalveolar lavage (BAL) (closed triangles, n=24) and control BAL (open triangles, n=4) are expressed as cells ×103/BAL sample. (B) Peripheral blood DC percentages in RSV (closed triangles, n=19) and control cases (open triangles, n=5). (C) CD83, CD40, ICOS-ligand, PDL-1 and CD80 expression on peripheral blood cDCs from RSV cases (closed symbols, n=19 (n=15 for CD80)) and controls (open symbols, n=5 (n=3 for CD83, n=4 for PDL-1)). Data are expressed as medians (IQR). p Values were determined by Mann-Whitney U tests.
Peripheral blood cDCs upregulate CD83 and CD40 expression in infants with RSV bronchiolitis
To determine the level of cDC activation, we assessed the expression of the activation marker CD83 and of the DC co-stimulatory molecules CD86, CD40, CD80, ICOS-ligand and PDL-1. In the peripheral blood, CD83 and CD40 were expressed on a significantly higher percentage of cDCs in infants with RSV bronchiolitis compared with controls. This also appeared to be the case for ICOS-ligand (4.4 (IQR 1–19)% vs 1.1 (IQR 0.5–1.7)%, p=0.08) and PDL-1 (3.2 (IQR 0–9.3)% vs 0 (IQR 0–0.15)%, p=0.08) (figure 4C), but these differences did not reach statistical significance.
On BAL cDCs, we did not find differences between RSV cases and controls in CD86 (3.1 (IQR 0.2–21.2)% vs 5 (IQR 0–28.4)%, p=0.61), CD80 (0 (IQR 0–2.7)% vs 3.7 (IQR 0–8.6)%, p=0.33), CD40 (7.6 (IQR 1.3–12.1)% vs 12.2 (IQR 0–14.8)%, p=0.87), but ICOS-ligand expression appeared to be higher (4.9 (IQR 0.9–17.5)% vs 0 (IQR 0–18.2)%, p=0.11).
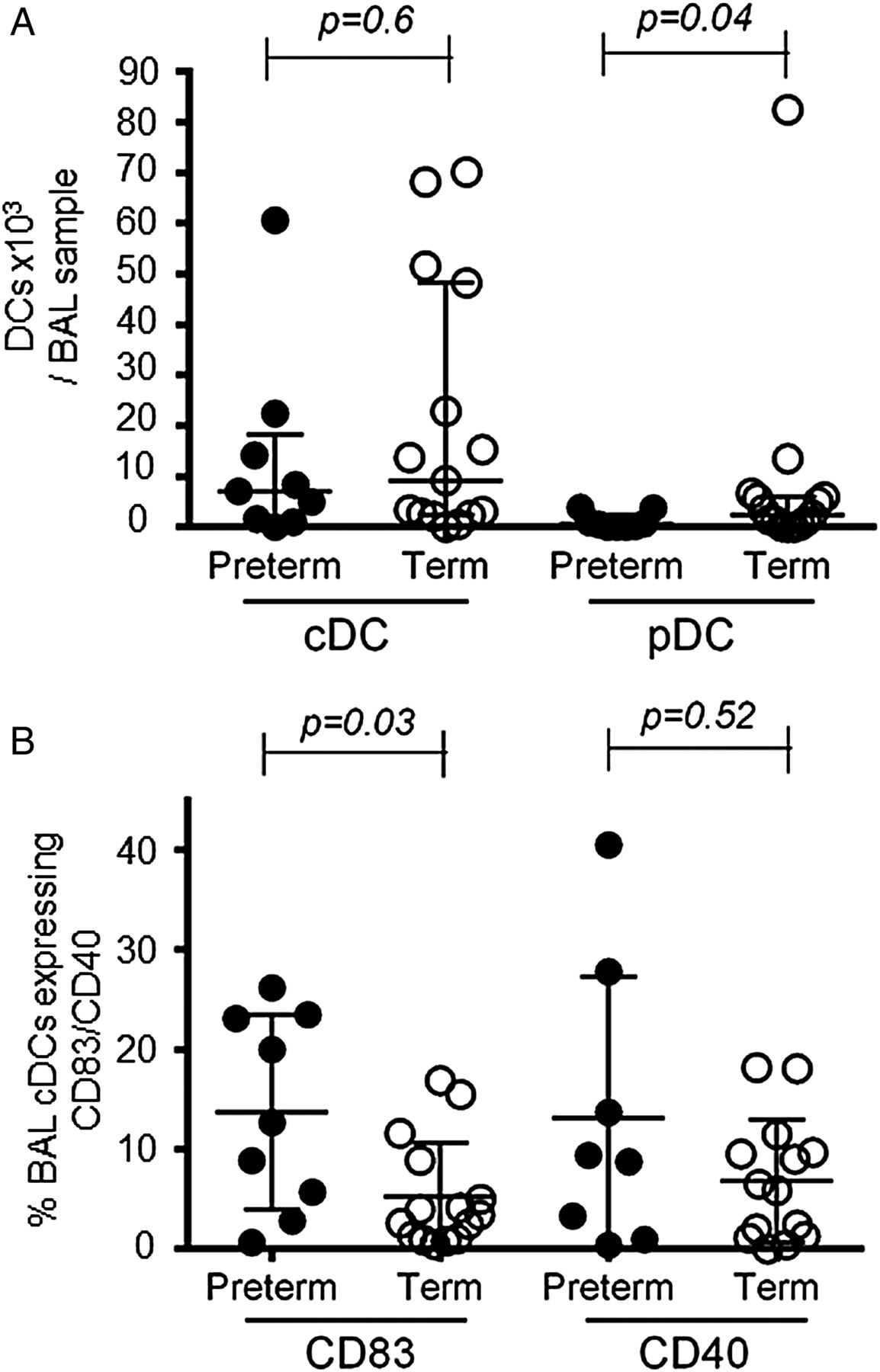
Preterm infants with RSV bronchiolitis have lower numbers of pDCs than term born infants and increased cDC activation marker expression
Next we asked whether there are differences in DC responses in RSV bronchiolitis between preterm and term born infants. We did not observe any difference in the percentage of BAL fluid recovered (39.5 (IQR 22.9–53.1)% vs 42.9 (IQR 20.6–48.2)%, p=0.89) or total BAL cellularity (1.5 (IQR 0.2–2.6) vs 1 (IQR 0.3–3.1) × 106 cells/BAL sample, p=0.87) between preterm and term born infants.
While there was no significant difference in BAL cDC percentages (0.4 (IQR 0.2–1.6)% vs 1.1 (IQR 0.5–1.3)%, p=0.34) or numbers (figure 5A) between preterm and term born infants, preterm infants expressed CD83 on a significantly larger percentage of BAL cDCs (figure 5B). No significant differences were found in CD40 (figure 5B), CD80, CD86, programmed death ligand (PDL)-1 or inducible T cell co-stimulator (ICOS)-ligand expression (additional data in online supplement).
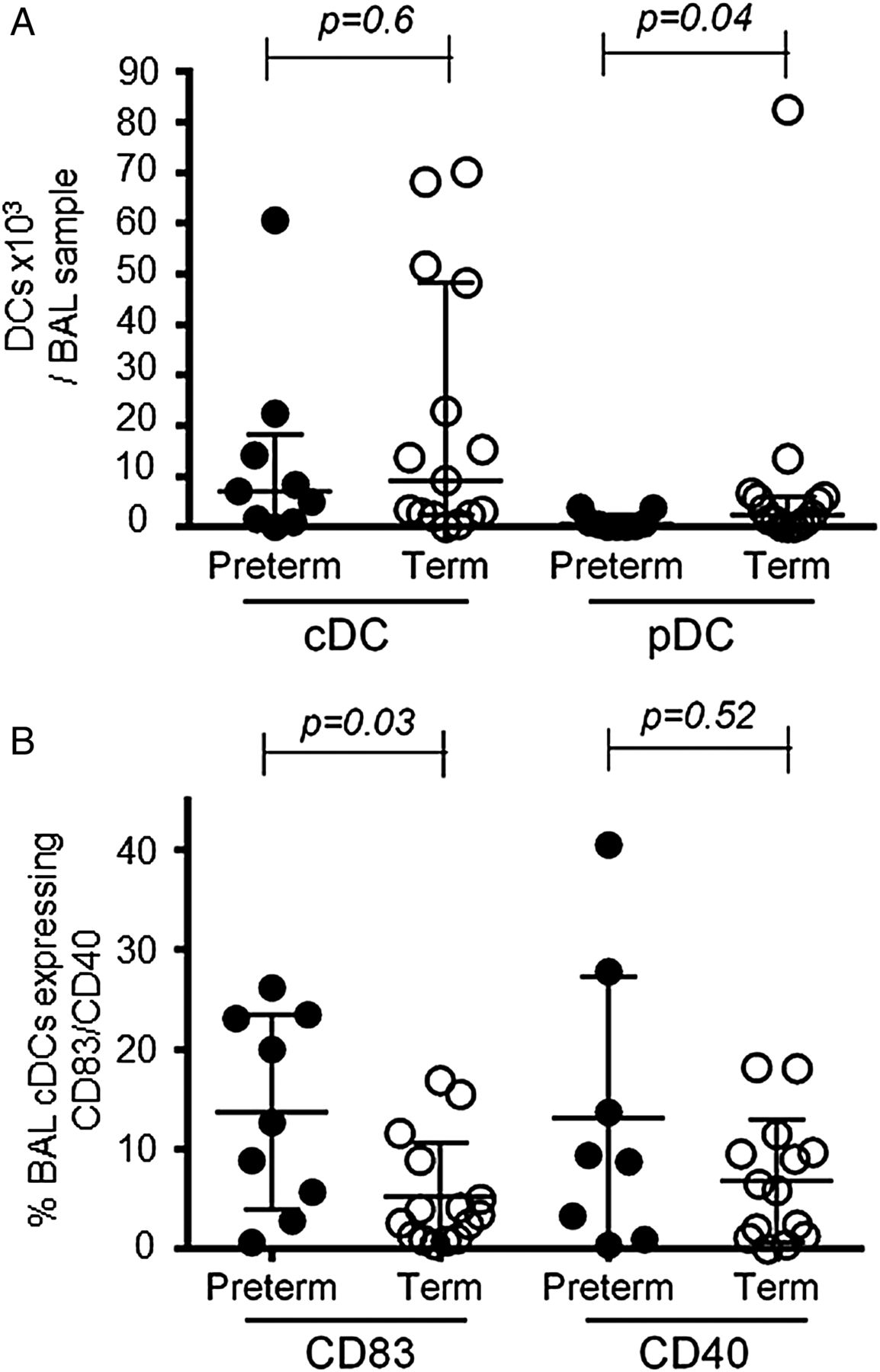
Plasmacytoid dendritic cell (pDC) numbers are lower and conventional dendritic cell (cDC) activation marker expression is increased in bronchoalveolar lavage (BAL) samples from preterm compared with term born infants with respiratory syncytial virus bronchiolitis. (A) BAL cDC and pDC numbers were compared between preterm (closed symbols, n=9) and term (open symbols, n=15) infants. Data are expressed as medians (IQR). p Values were determined by Mann-Whitney U tests. (B) Expression of CD83 and CD40 on cDCs in BAL samples from preterm (closed symbols, n=9 (n=8 for CD40)) and term born infants (open symbols, n=15 (n=14 for CD40)). Data are expressed as medians (IQR). p Values were determined by Mann-Whitney U tests.
In contrast to cDCs, BAL-pDC numbers were significantly lower in preterm compared with term born infants (figure 5A).
In the peripheral blood, percentages of cDCs did not differ between the two groups (additional data in online supplement), whereas pDC percentages were significantly lower in those born at term (0.2 (IQR 0.1–0.2)% vs 0.1 (IQR 0.01–0.1)%, p=0.02).
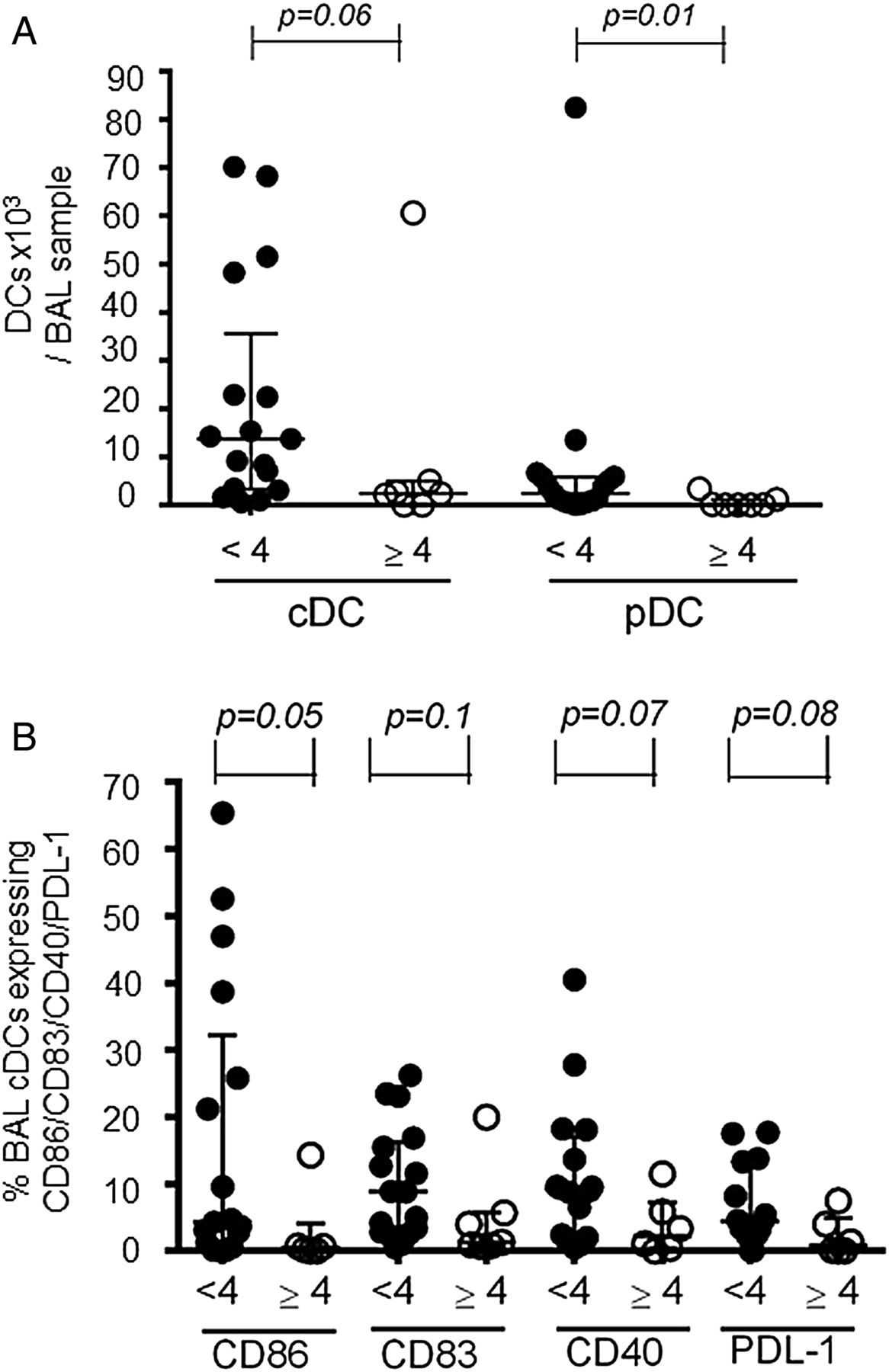
Older infants with RSV bronchiolitis have lower airway pDC numbers
Given that the highest risk of severe RSV bronchiolitis is in infants aged 2–4 months,8 we stratified the RSV cases into younger (<4 months of age) and older (≥4 months of age) infants. Comparing DC responses, we found that the numbers of BAL pDCs were significantly higher in younger than in older infants (figure 6A) and correlated negatively with increasing age (R −0.58 (95% CI −0.8 to −0.2), p=0.001, online supplementary figure S2). Furthermore, the percentage of pDCs in the peripheral blood was higher in younger compared with older infants (0.16 (IQR 0.1–0.24)% vs 0.07 (IQR 0.03–0.15)%, p=0.07) without reaching statistical significance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Older infants with respiratory syncytial virus bronchiolitis have fewer bronchoalveolar lavage (BAL) plasmacytoid dendritic cells (pDCs), while younger infants may have more activated BAL conventional dendritic cells (cDCs). (A) BAL cDC and pDC numbers were compared between infants <4 months of age (closed symbols, n=17) and infants ≥4 months of age (open symbols, n=7). (B) Expression of CD86, CD83, CD40 and PDL-1 on BAL cDCs was compared between infants aged <4 months (closed symbols, n=17 (n=16 for CD40 and n=15 for PDL-1)) and ≥4 months (open symbols, n=6 (n=7 for CD83)). Data are expressed as median (IQR). p Values were determined by Mann-Whitney U tests.
When assessing BAL cDCs, their numbers appeared to be higher in younger than in older infants (figure 6A), as did the percentages of BAL cDCs expressing CD86 (4.3 (IQR 0.9–32.3)% vs 0.4 (IQR 0–4.1)%, p=0.05), CD83 (8.9 (IQR 2.7–16.2)% vs 1.3 (IQR 0.8–5.7)%, p=0.09), CD40 9.2 (IQR 2.1–17)% vs 2.2 (IQR 0.3–7.3)%, p=0.07) and PDL-1 (4.5 (IQR 2.4–13.3)% vs 0.8 (IQR 0–4.9)%, p=0.08) (figure 6B). However, these differences did not reach statistical significance. There was no difference in ICOS-ligand expression (additional data in online supplement).
In the peripheral blood, the percentage of cDCs did not differ significantly between younger and older infants (0.13 (IQR 0.03–0.4)% vs 0.1 (IQR 0.02–0.1)%, p=0.12).
BAL and serum cytokines in RSV bronchiolitis
In the BAL of RSV cases, the T cell cytokine IL-2, Th1 cytokine IFNγ, Th2 cytokines IL-13 and IL-10, and pro-inflammatory cytokines IL-1β, IL-6, IL-8 and TNF-α were all significantly higher than in controls (table 1). Serum concentrations of IFNγ, IL-10, IL-6 and TNF-α were also significantly higher in RSV bronchiolitis compared with controls.
Bronchoalveolar lavage (BAL) and serum concentrations in infants with respiratory syncytial virus (RSV) bronchiolitis and controls
When comparing cytokines between younger and older infants, we found higher BAL IFNγ levels (44.4 (IQR 18.3–60.5) vs 10.7 (IQR 3.9–19.1) pg/mL, p=0.02) and higher serum TNF-α levels (2.3 (IQR 1.8–2.8) vs 1.4(IQR 1.1–1.9) pg/mL, p=0.002) respectively, without differences in any of the other cytokines measured. Comparing preterm and term born infants, there were no differences in BAL cytokines, but preterm infants had higher serum levels of IL-2 (0.3 (IQR 0.2–0.4) vs 0.1 (IQR 0.03–0.3) pg/mL, p=0.03) and IL-1β (0.2 (IQR 0.1–0.3) vs 0.1 (IQR 0.06–0.2) pg/mL, p=0.04).
Interestingly, the concentrations of the innate pro-inflammatory cytokines IL-6, TNF-α and IL-8, and of the T-cell-derived cytokines IL-13, IL-10, IL-2 and IL-4 correlated with BAL cDC numbers and IL-10 concentrations also correlated with pDC numbers (see online supplementary figure S3). Correlations between other immune cells and cytokines in BAL are shown in online supplementary table S1.
Discussion
Data on lower airway DC responses in infants with RSV bronchiolitis is lacking. Here, we determined, to our knowledge for the first time, the presence and phenotype of DCs and associated lymphocytes in the lower airways of infants with RSV bronchiolitis to help elucidate their role in this disease.
We show significant increases in numbers of BAL cDCs in infants with RSV bronchiolitis compared with healthy controls. This parallels findings of increased cDC numbers in the nasal mucosa of children with RSV infection,18 ,20 and our previous observation of increased lung cDCs in the mouse model of RSV infection,5 ,6 further validating this model. The origin of the lower airway DC during RSV bronchiolitis is not clear. DCs or their precursors may be recruited into the lung from the peripheral blood and bone marrow. Our and other investigators' observations of significantly decreased percentages of peripheral blood cDCs and pDCs in RSV cases compared with controls18 may support this notion. However, cDC populations could also expand in the lung from local precursors, as we previously observed in RSV-infected mice.21
Lung cDCs have an activated, pro-inflammatory phenotype in RSV-infected mice.5 ,21 Similarly, we found increased expression of the cDC activation marker, CD83 and the co-stimulatory molecule, CD40 on peripheral blood cDCs of infants with RSV bronchiolitis compared with controls. We did not however find such difference between RSV cases and controls on BAL cDCs. This could be explained if the controls that provided adequate BAL samples for flow cytometry were recovering from a subclinical or minor respiratory infection, which was not recognised at recruitment, a realistic possibility given that these samples had a higher cellularity than the majority of controls and robust neutrophil and lymphocyte populations.
Activated cDCs can induce primary T cell responses and determine their nature and T cells, in turn, have a central role in the development of RSV-induced inflammation and disease in mouse models.3 ,4 However, their role in infants with RSV bronchiolitis is unclear. In fatal RSV cases T cells may be absent22 or occur only at very low frequencies.23 In contrast, in non-fatal RSV bronchiolitis increased BAL CD4+ and CD8+ T cell numbers24 and the presence of lung tissue T cells25 have been reported. Here, we found reproducible CD4+ and CD8+ T cell populations and an apparently increased CD8+/CD4+ T cell ratio in the lower airways of infants with RSV bronchiolitis, but did not detect statistically significant differences compared with controls.
Stimulated cDCs can also activate natural killer (NK) and NK T cells.26–29 We describe for the first time, to our knowledge, the substantial accumulation of NK and NK T cell populations in the lower airways of infants with RSV bronchiolitis. This is in contrast to previous reports that found <1% of BAL cells to be NK cells and only rare staining of CD56+ NK cells in lung tissue from infants with fatal RSV infection.22 ,24 However, our findings are in agreement with the mouse model, where activated NK cells accumulate in the lung early in RSV infection.30 ,31 Although we did not assess the activation status of NK and NK T cells, we speculate that high BAL cDC numbers in RSV bronchiolitis, in addition to macrophages,32 may contribute to NK and NK T cell activation, and thus possibly to lung immune injury, as seen in RSV infection in mice.31
Previous studies have consistently found neutrophils to be the predominant leucocyte population in RSV bronchiolitis, accounting for ∼76% and ∼93% of cells in the lower and upper airways, respectively.33 ,34 Monocytes have also been described in RSV-BAL, along with strong expression of the CD16 macrophage antigen in lung tissue samples from infants with fatal RSV infection.24 ,25 We confirm the accumulation of neutrophils, as the predominant cell type, and of monocytes and macrophages in the lower airways in RSV bronchiolitis.
Next we asked whether specific DC response patterns are associated with individual risk factors for severe RSV bronchiolitis. Premature birth is a risk factor for hospitalisation and longer duration of mechanical ventilation in RSV infection.9 ,10 We observed significantly lower BAL pDC numbers in preterm compared with term born infants with RSV bronchiolitis. Based on findings in the mouse models, recruitment of antiviral IFNα-producing pDCs to the lung is a normal early response to RSV infection, required to limit viral load.6 ,7 If this response is inadequate in preterm infants, this may lead to increased RSV load, resulting in more severe disease.35 ,36 Interestingly, the activation markers CD83 and CD40 were expressed on more BAL cDCs in preterm infants. This could also be linked to the low pDC response. In addition to their antiviral activity, activated pDCs are known to produce IL-10 and are thought to have regulatory anti-inflammatory properties in RSV infection7 and could potentially limit cDC activation.37 The correlation of pDCs and IL-10 that we observed in the BAL of RSV cases supports this notion.
The other major group at risk of severe RSV bronchiolitis are infants of 2–4 months of age.8 We therefore stratified RSV cases into ‘younger’ (<4 months) and ‘older’ infants (≥4 months) and found significantly lower BAL-pDC numbers in older infants and a negative correlation of these cells with postnatal age. As discussed above for preterm infants, these findings may indicate both inadequate regulation of inflammation and an inadequate antiviral response during RSV infection. However, RSV load was not assessed in this study, leaving us unable to determine any correlation with pDC numbers.
When assessing BAL-cDCs, their numbers and expression of co-stimulatory molecules and an activation marker appeared to be higher in younger than in older infants. Although there was no difference in BAL cytokines between younger and older infants, concentrations of innate pro-inflammatory (IL-6, TNF-α and IL-8) and T cell cytokines (IL-13, IL-10 and IL-2) correlated with BAL cDC numbers. It therefore seems possible that innate pro-inflammatory cytokines activate airway cDCs, which in turn may drive T cell responses, contributing to an intense inflammatory response to RSV infection in younger infants. Although we do not have clinical data on severity, stage of disease or duration of illness, these cytokine data support the notion of a link between BAL cDC numbers and severity of inflammation and disease.
Based on these observations, we speculate that lower airway DC response patterns may allow the definition of different endotypes of RSV bronchiolitis; one with a strong, pro-inflammatory cDC response leading to excessive pulmonary inflammation, and another with insufficient viral control associated with a diminished antiviral pDC response. Being able to recognise such endotypes would enable targeting of future antiviral and anti-inflammatory therapy to the appropriate patients with bronchiolitis.
Acknowledgments
The authors thank Drs Sarah Howie and Donald Davidson for helpful discussion of the manuscript. They also thank all parents/carers who agreed for their children to be enrolled in this study.
References
Footnotes
Contributors Concept design and ethics: PF, RL, KMacK, UT and JS. Analysis and interpretation: AK, PF, HMcS, UT, JS, SW and AB. Drafting of manuscript: AK, RL, UT and JS. Patient recruitment and sampling: DK, LW, KR, JMcC, FM, RL, UT and JS.
Funding This work was supported by the Chief Scientist Office Scotland (ETM-108), the Medical Research Council (MR-K002589-1) and the Wellcome Trust (067454/Z/02/c).
Competing interests None declared
Patient consent Parental/guardian written consent obtained.
Ethics approval The study was approved by the South East Scotland Research Ethics Committee 03 (08/S1103/50).
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Authors have full access to all of the data and take responsibility for the integrity and the accuracy of the data and analysis.