Article Text

Abstract
Rationale Aminoglycoside (AG) resistance of Pseudomonas aeruginosa in cystic fibrosis (CF) is associated with poorer clinical outcomes and is usually due to overexpression of the efflux pump MexXY. MexXY is regulated by mexZ, one of the most commonly mutated genes in CF P. aeruginosa isolates. Little is known about the evolutionary relationship between AG resistance, MexXY expression and mexZ mutations.
Objectives To test the hypothesis that AG resistance in P. aeruginosa develops in parallel with higher MexXY expression and mexZ mutations.
Methods CF P. aeruginosa isolates were compared for chronically infected (CI) adults, CI children and children with new infection.
Measurements One P. aeruginosa isolate from each patient was analysed for mexZ mutations, mexY mRNA expression and amikacin resistance.
Main results 56 patients with CF were enrolled: 21 children with new P. aeruginosa infection, 18 CI children and 17 CI adults. Amikacin resistance and mexY mRNA expression were higher in cohorts with longer P. aeruginosa infection. The prevalence of non-conservative mexZ mutations was 0%, 33% and 65% in children with new infection, CI children and CI adults, respectively. The same trend was seen in the ratio of non-conservative to non-synonymous mexZ mutations. Of isolates with non-conservative mexZ mutations, 59% were amikacin-resistant compared with 18% of isolates with non-synonymous mutations. The doubling rate of amikacin resistance and non-conservative mexZ mutations was approximately 5 years.
Conclusions P. aeruginosa mexZ mutations undergo positive selection resulting in increased mexY mRNA expression and amikacin resistance and likely play a role in bacterial adaption in the CF lung.
- Cystic Fibrosis
- Respiratory Infection
- Bacterial Infection
- Bronchiectasis
Statistics from Altmetric.com
Key messages
What is the key question?
Given the worse prognosis for patients with cystic fibrosis (CF) having antibiotic-resistant Pseudomonas aeruginosa, what is the mechanism of aminoglycoside (AG) resistance in P. aeruginosa in CF?
What is the bottom line?
Mutations in the mexZ gene, a regulator of the MexXY multidrug efflux pump, increase and become fixed over time, leading to increased MexXY expression and AG resistance.
Why read on?
Understanding the mechanism by which P. aeruginosa develops AG resistance may provide a more rational approach to antibiotic stewardship in order to delay chronic P. aeruginosa infection and minimise antibiotic resistance in CF.
Introduction
Cystic fibrosis (CF) occurs in approximately 1 in 3500 newborns1 and is caused by mutations in the CF transmembrane regulator (CFTR) gene, which can lead to abnormal function in multiple organs; in the lung, CFTR mutations lead to bronchiectasis.2 Development of bronchiectasis is caused by a vicious cycle of infection, inflammation and mucus obstruction, with recent data suggesting that infection may be the antecedent event.3
One of the most important infectious agents in CF is Pseudomonas aeruginosa, which affects approximately 80% of patients with CF and is associated with severe pulmonary exacerbations, accelerated loss of lung function and increased mortality.4 ,5 P. aeruginosa infection early in the course of CF phenotypically resembles acute non-CF infections but evolves genotypically and phenotypically to cause chronic infection in the lung. The mechanisms by which P. aeruginosa adapts to the CF airway to maintain prolonged residence are incompletely understood but are related in part to its very large genome and the ease with which it can mutate.6 These properties facilitate selection of adaptive mutations, of which the most important is the development of antibiotic resistance, which may facilitate prolonged lung residence and contribute to poorer outcomes in patients with CF.7 ,8
The aminoglycosides (AGs) are integral to the treatment of P. aeruginosa; however, resistance to these antibiotics often develops due to increased expression of the MexXY-OprM efflux pump.9–11 The MexXY-OprM system comprises a cytoplasmic membrane antibiotic-proton antiporter (MexY), an outer membrane porin (OprM) and a periplasmic membrane fusion protein, which joins the membrane-associated components together (MexX).12 MexXY has multiple functions, including the expulsion of antibiotics. While wild-type P. aeruginosa has constitutively low MexXY levels, elevated MexXY has been detected in AG-resistant P. aeruginosa strains in patients with CF and patients without CF.10 ,13–16 The MexXY components are encoded by an operon under the control of an adjacent repressor gene, mexZ. MexZ protein contains a DNA-binding domain (DBD) and a ligand-binding C-terminal domain (CTD) connected by a linker. The MexZ protein dimerises via the CTD domain and directly binds a specific site of the mexZ-mexX intergenic DNA via the DBD to repress transcription of the mexXY operon.17 On binding of ligand to CTD, the DBD changes conformation and releases the protein from the DNA to allow MexXY expression.18 Importantly, the mexZ gene is one of the most frequently mutated genes in CF P. aeruginosa isolates.19–21 However, there are little data on the relationship between mexZ mutations and AG resistance.10 ,15 In addition, the type and locations of mexZ mutations and their relationship to MexXY expression and AG resistance have yet to be characterised.
We hypothesised that the prevalence of functionally important P. aeruginosa mexZ mutations would increase with residence time in the CF lung and be associated with increased MexY mRNA expression and AG resistance.
Materials and methods
Subjects and definitions
Sample collection and study analysis was approved through the institutional review boards at both Northwestern University and Lurie Children's Hospital of Chicago. Informed consent was obtained from all participants or parents prior to sample collection. Written assent was obtained for participating children aged ≥12 and <18 years. Patients were considered adults if they were at least 18 years of age at the time of enrolment. Subjects with a confirmed CF diagnosis (positive sweat chloride and/or two disease-causing CFTR mutations) and a respiratory culture positive for P. aeruginosa were studied between the years 2003 and 2006. The study design was cross-sectional, in which each patient was assigned to one of the three cohorts and from whom a single P. aeruginosa isolate was used. Patients were considered chronically infected (CI) if they had at least one P. aeruginosa-positive respiratory culture in the 2 years preceding the index culture. Patients who had never had P. aeruginosa or had not grown it in the 2 years preceding study enrollment were considered to have a new infection.
For CI patients, respiratory specimens were collected every 6 months for approximately 2 years. For patients without prior P. aeruginosa isolates, surveillance cultures were obtained every 3–6 months and isolates were studied at time of the first P. aeruginosa-positive culture. Spirometry was performed at each visit and used FEV1 as a measure of pulmonary function. The clinical characteristics and year of first P. aeruginosa infection was ascertained from the medical record or Cystic Fibrosis Foundation (CFF) registry. Duration of P. aeruginosa infection was calculated as the difference in time between first infection and enrolment in the study, with newly infected patients being assigned a duration of zero.
Collection and processing of bacterial isolates
P. aeruginosa clinical isolates used in this study have been previously reported.22 Respiratory samples (sputum, oropharyngeal swabs or bronchoalveolar lavage) were processed by microbiology laboratories at Northwestern Memorial Hospital and Lurie Children's Hospital and P. aeruginosa was identified using criteria approved by the National Committee for Clinical Laboratory Standards. P. aeruginosa isolates were plated and a single isolate was grown in Luria-Bertani broth at 37°C and 250 rpm until log phase growth was achieved. Simultaneous aliquots were then separated for measurement of RNA, DNA sequencing and clinical antibiotic susceptibility testing, as described below. This ensured that each assay was performed on isolates grown under identical conditions.
RNA isolation and complementary DNA confirmation
Bacterial lysis was achieved by physical agitation and chemical disruption using lysozyme and proteinase K. RNA was obtained with RNA Protect Bacteria Reagent (Qiagen, Valencia, California, USA) and isolated using column extraction with the RNeasy Mini Kit (Qiagen). DNase digestion was performed and RNA was quantified using a NanoDrop 2000 ultraviolet spectrophotometer (Thermo Scientific, Wilmington, Delaware, USA). Reverse transcription (RT) PCR was performed using the Advantage RT-for-PCR Kit (Clonetech, Mountain View, California, USA) with initial heating to 70°C followed by incubation at 42°C for 1 hour and denaturation at 94°C for 5 min with subsequent cooling on ice. Primers used in this study are listed in online supplementary table S2 and unique primers for this study were designed with the aid of the PrimerBLAST programme (http://www.ncbi.nml.nih.gov/tools/primer-blast/). Standard PCR was used with both complementary DNA (cDNA) and RNA as template with primers for mexY and rspL as previously published to identify the product of appropriate size and to ensure the absence of genomic DNA contamination.13 ,23
Supplemental material
Real-time quantitative PCR
There is a strong correlation between mexX and mexY mRNA expression and the MexXY protein is predominantly transcriptionally regulated. Thus, we opted to analyse mexY mRNA expression as a marker for MexXY protein expression. Real-time RT-quantitative PCR was carried out in triplicate using SYBR green supermix (Bio-Rad, Hercules, California, USA) and cDNA as template. cDNA was amplified using the iCycler iQ system (Bio-Rad). Cycle threshold values were normalised for amplification of housekeeping gene rspL.24 The concentration of primers and cDNA were 300 and 200 nM, respectively. For the PCR reaction, 2 μL of cDNA at 1:1 and 1:4 concentrations for the mexY and rspL were used, respectively. Standard 1:4 dilutions were established for each reaction using a stock solution of PAO1 cDNA to ensure consistency across samples. Expression levels were determined by using the average expression rate of triplicate compared with PAO1 (obtained from University of Washington) at the time of each reaction. Isolates were categorised as ‘hyperexpressors’ if they exceeded 25 times the expression rate of PAO1 based on dichotomous grouping of all isolates around the group median. Control strains used in this work (PAO3579, mexXY overproducer) and PAO280 (PAO3579 with deletion of mexXY) were generously provided by Herbert Schweizer and previously characterised.25
Traditional DNA sequencing
MexZ sequencing was performed in a two-stage process (sequence and gel extraction) due to low abundance of the mRNA. Aliquots reserved for DNA were pelleted and DNA was extracted from bacterial cells using free-thaw techniques and column extracted using the DNeasy spin columns (Qiagen). PCR was performed using primers MexZ2/MexZ2b to flank the gene of interest run on a 1% agarose gel for extraction. Excised bands were prepared for sequencing with the QIaquick gel extraction kit (Qiagen) and eluted in water. Traditional DNA sequencing was then performed through the Northwestern University Genomics Core Facility starting with 10 ng of template DNA. The sequencing reaction was conducted with BigDye chemistry in an Applied Biosystems (ABI) GeneAmp 9700 thermocycler (Thermo Fisher Scientific, Waltham, Massachusetts, USA), followed by clean-up using Edge Bio Performa V3 silica gel plates. An ABI 48-capillary 3730 DNA analyser was then used for electrophoresis, sequence generation and final reporting. MexZ sequences were compared with PAO1 using nucleotide BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) to detect all base pair mutations. Nucleotide sequences were then converted to protein sequence (CLC Sequence Viewer 7.6.1, CLC bio, Denmark) and again compared with PAO1 to determine the presence of non-synonymous mutations. Non-synonymous mutations were defined as mutations which led to a different amino acid (AA) sequence. Non-conservative and conservative mutations were defined as non-synonymous mutations that led to a change or no change, respectively, in the predicted protein function, based on the Sorting Intolerant From Tolerant (SIFT) programme as previously described.26
AG susceptibility testing
The final aliquot of each log phase isolate was tested by clinical microbiology lab at Lurie Children's for antibiotic susceptibility using the Kirby Bauer method per standard clinical practice for CF P. aeruginosa samples. All testing was performed using Clinical and Laboratory Standards Institute methods and reported as sensitive, intermediate or resistant. Isolates reported as intermediate were considered to be resistant for the purposes of this study.
Statistics
The subjects were grouped into three cohorts in order to facilitate the analysis for binary variables such as mexZ mutations and amikacin resistance. The data were analysed using Prism 4 (GraphPad Software, La Jolla, California, USA). All data were displayed as means±SE or medians±range as appropriate. The association between duration of infection and mexY mRNA expression was examined using Spearman's rank correlation coefficient. Statistical significance was determined by unpaired t tests, one-way analysis of variance and χ2 analysis as appropriate. A p value of <0.05 was considered statistically significant.
Results
Demographics
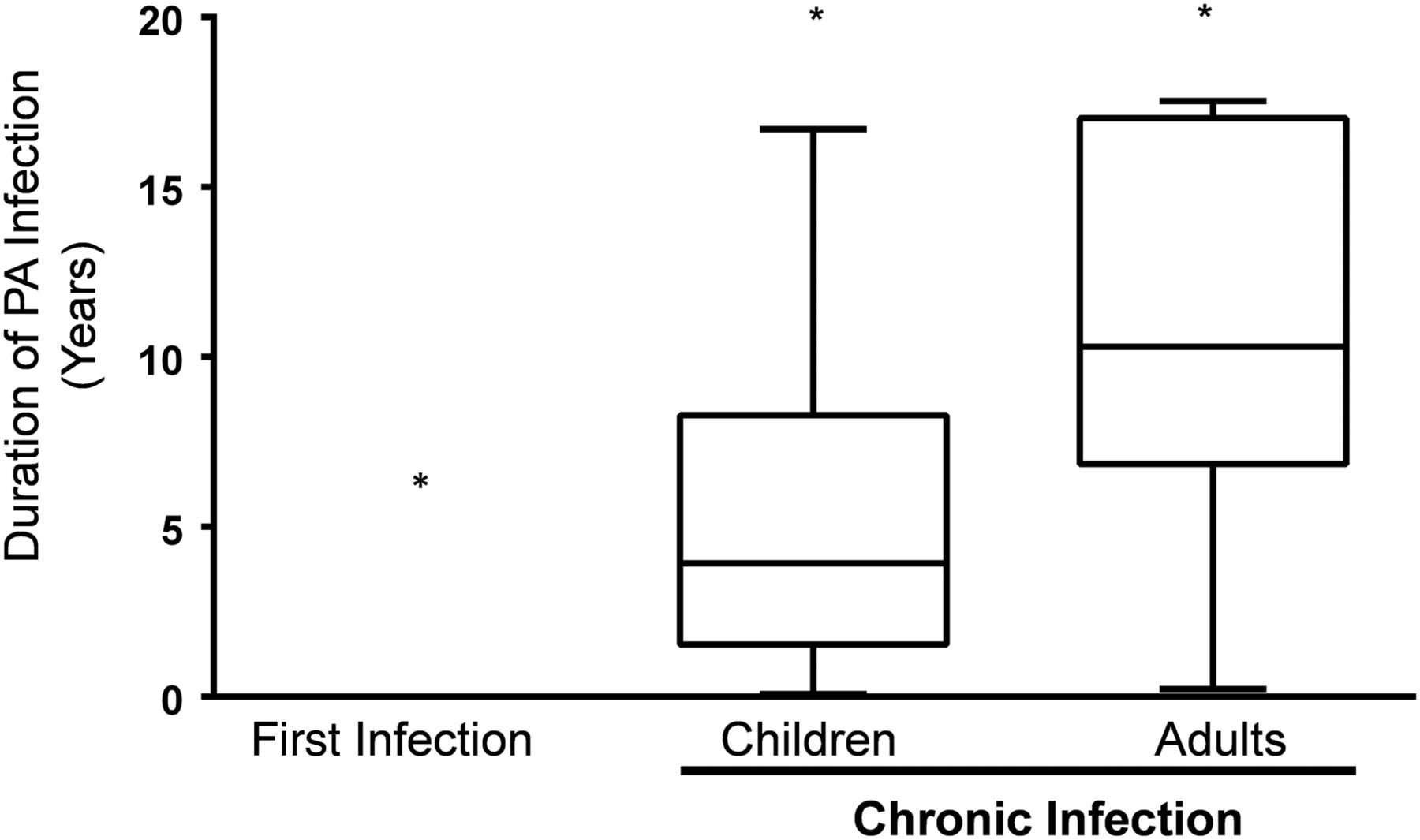
Fifty-six patients with CF (39 children <18 years of age and 17 adults) were included in the study. Selected demographics are shown in table 1. Eighteen (46%) children and all 17 adults were CI with P. aeruginosa. The remaining 21 children were enrolled after culturing P. aeruginosa for the first time (table 1). The mean age of the new P. aeruginosa infection cohort was lower (p<0.01) compared with CI children and adults. There was no statistical difference in the proportion of homozygous delF508 patients across the three cohorts. The duration of P. aeruginosa infection was 11.2±5.2 and 5.8±4.8 years for the CI adult and children cohorts, respectively (p<0.002; figure 1).
Selected demographics of study cohorts
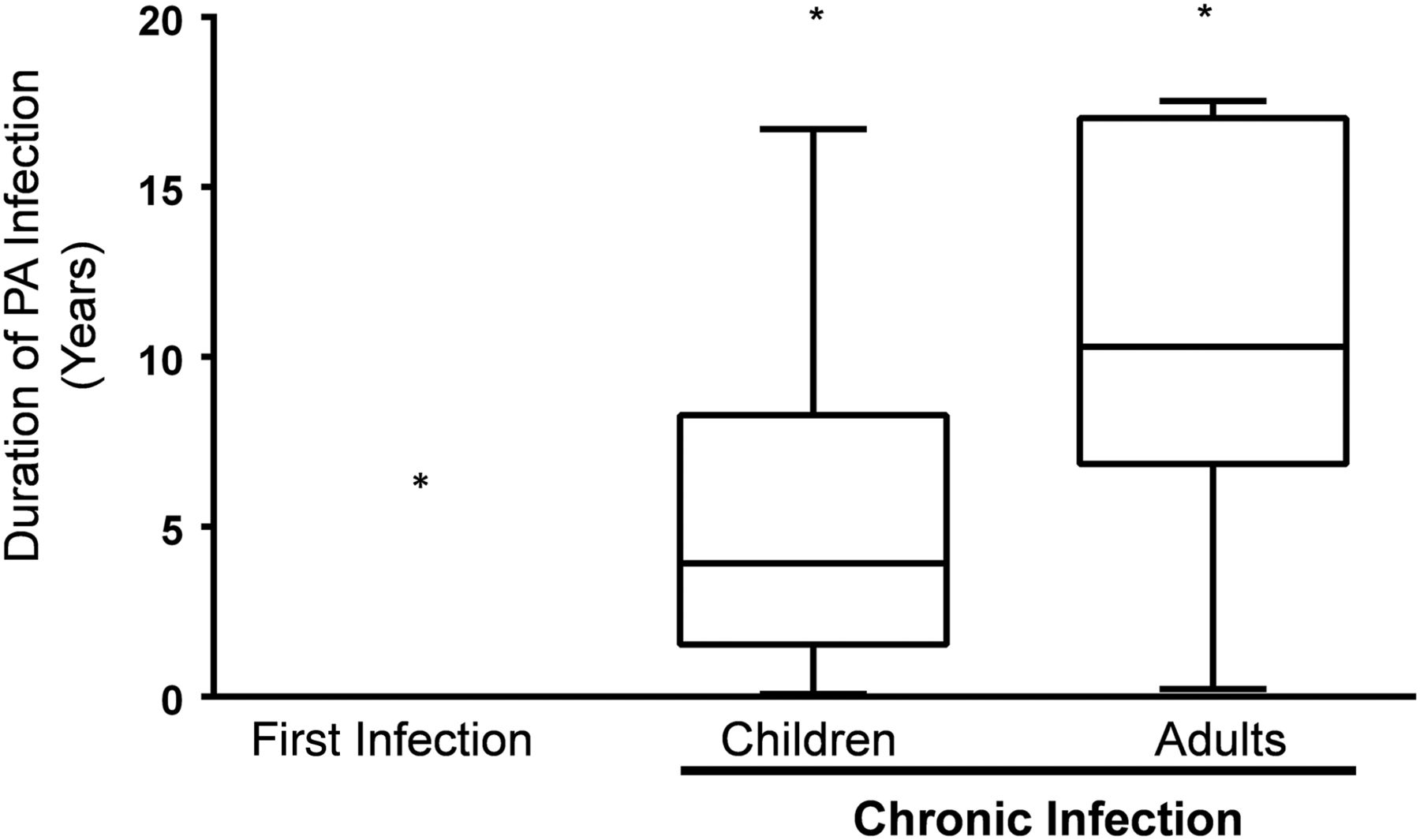
Duration of Pseudomonas aeruginosa infection was significantly different between the three cohorts. Mean duration of P. aeruginosa infection in the newly infected, chronically infected (CI) children and CI adult cohorts are shown (*p<0.001 for comparison of means of each cohort).
Mexz mutations
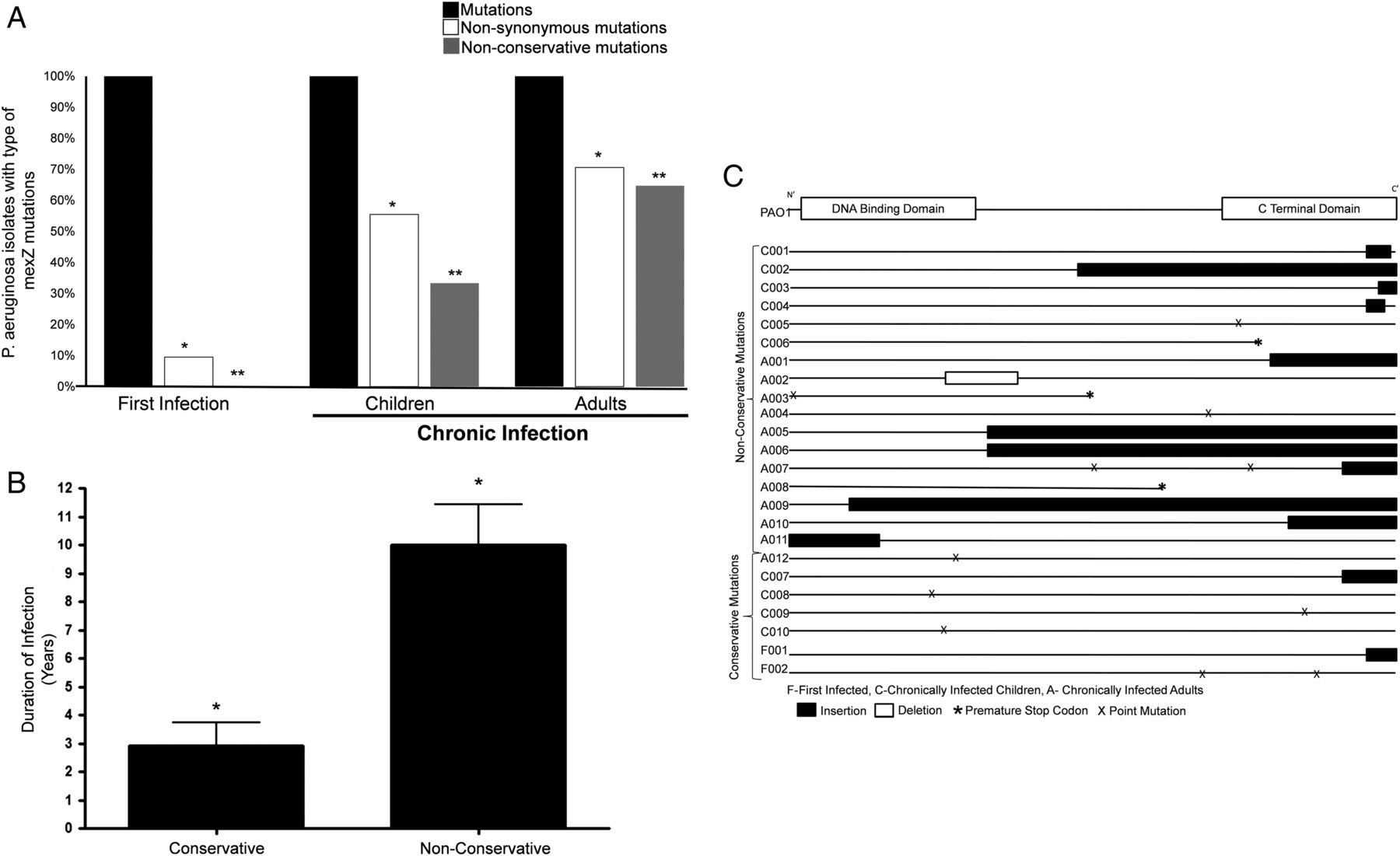
The rate at which mexZ mutations develop in CF P. aeruginosa isolates and whether they impact protein function (ie, how frequent non-conservative mutations occur) have not been reported. Based on the recently solved three-dimensional structure of MexZ, there are three components of the protein that are felt to be important for optimal function.18 ,27 To exclude the possibility that the referent PAO1 strain carries a mutation in mexZ, we compared its sequence with strains obtained from three independent laboratories and found them to have an identical sequence (data not shown). Next, to assess the relationship between P. aeruginosa residence time and the functional consequences of mexZ gene mutations, we sequenced mexZ of each P. aeruginosa isolate. In the newly infected cohort, all isolates had unique mexZ sequences that differed from PAO1 and from each other, of which 2 of 21 (9.5%) resulted in a non-synonymous mutation (ie, a base pair change resulting in an AA change). The two missense mutations were unique and conservative (ie, not predicted to affect MexZ protein function; figure 2A). In comparison, isolates from all of the CI children had unique mexZ sequences that differed from PAO1 and from each other, of which 10 of 18 (56%) led to a change in the AA sequence. Of these 10 isolates, 5 had insertion/deletions and 5 had non-synonymous single-nucleotide substitutions (figure 2A, C). Six of these sequence changes were predicted to change protein function, including all isolates with insertions/deletions. In CI adults, all of the isolates had a different mexZ sequence from PAO1, of which 12 (76%) led to a change in AA sequence. Of these 12 isolates, 11 (65%) contained insertion/deletions or premature stop codons that were predicted to alter protein function (figure 2A, C, exact sequences reported in online supplementary figure S1). Of the 17 isolates with non-conservative mutations, 12 were predicted to alter the CTD of MexZ, 2 were in the DBD and 3 had premature termination codons predicted to produce no functional protein. The prevalence of both non-synonymous and non-conservative mutations in the isolates increased as the residence time increased across cohorts (p<0.001; figure 2A). Further, the proportion of non-synonymous mutations that were non-conservative was 0% in the newly infected group, 60% in the CI children and 92% in the CI adults.

In cystic fibrosis Pseudomonas aeruginosa infection, mexZ mutations increase with time and are positively selected primarily due to large deletions and insertions. (A) The mexZ gene for each P. aeruginosa isolate was sequenced and compared with the reference PAO1 strain and analysed to determine if sequence changes were non-synonymous (ie, changed amino acid sequence) and non-conservative (ie, changed predicted protein function). Percentage of P. aeruginosa isolates with total, non-synonymous and non-conservative mexZ mutations in from each cohort is shown (*, # p<0.01 for differences in percentage for both non-synonymous and non-conservative mutations). (B) Mean duration of infection for P. aeruginosa isolates that carried conserved mexZ mutations compared with isolates with non-conserved mutations are shown (*p<0.001 for comparison of means). (C) Sites on the mexZ gene of each of the P. aeruginosa isolates with non-synonymous mutations, both with conserved and non-conserved protein function are shown.
The average residence time in the lung for all P. aeruginosa isolates with conserved mexZ mutations was 2.9 years and with non-conserved mexZ mutations was 10.0 years (p<0.001; figure 2B). Based on the mexZ mutation prevalence and average P. aeruginosa residence time in each cohort, we estimated that the prevalence of non-conservative mexZ mutations doubled approximately every 5 years of residence time in the CF lung.
We then assessed each non-conservative mexZ mutation with respect to the impact of its gene location on protein function. All but two of the non-conservative sequence changes were due to large insertions/deletions or premature stop codons. The two isolates with non-conservative missense mutations created new AA in the C-terminal ligand binding part of the protein. Further, all non-synonymous sequence changes were unique except for two isolates that had identical insertions. Interestingly, those two isolates came from a husband-wife couple with CF who were CI with P. aeruginosa.
Mexy mRNA expression
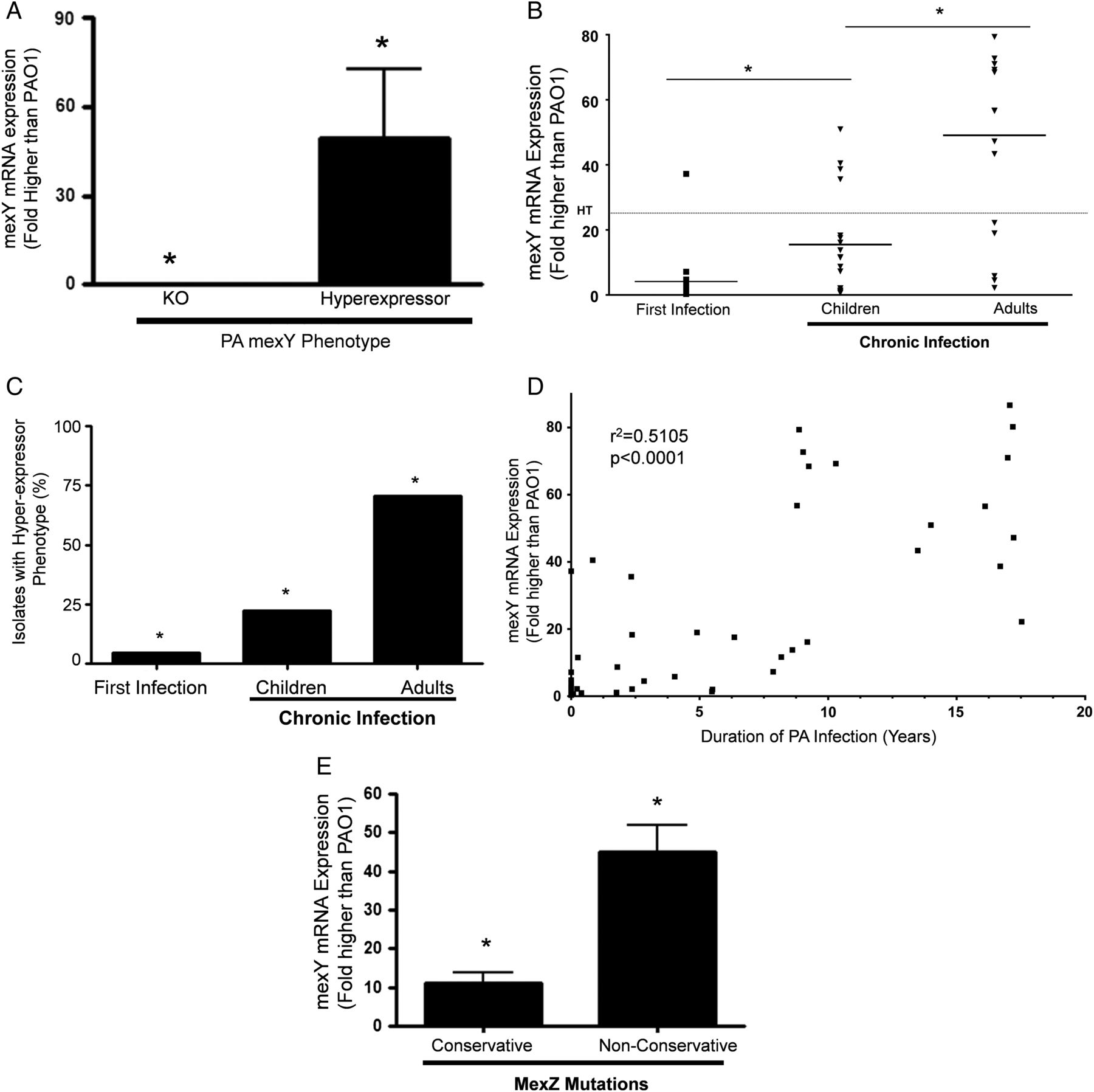
We used P. aeruginosa strains with mexY alleles causing a loss of expression or hyperexpression of mexY mRNA as negative and positive controls, respectively (figure 3A). MexY mRNA expression in each patient isolate was compared with a control P. aeruginosa strain, PAO1. Although expression levels were highly variable, median expression of mexY mRNA was highest in P. aeruginosa isolates from CI adults; the lowest expression was in the newly infected cohort and CI children had an intermediate level of mexY mRNA expression (p<0.001; figure 3B). Using a value of 25-fold higher expression than PAO1 as the definition of a ‘hyperexpressor strain’, the prevalence of hyperexpressor strains was higher in the CI adults (12/17; 70.6%) compared with CI children (4/18; 22.2%) and newly infected children (1/21; 4.7%) (p<0.001; figure 3C). When P. aeruginosa isolates from each patient were analysed with respect to duration of P. aeruginosa infection, there was a strong correlation between mexY mRNA expression and residence time in the CF lung (r2=0.51; p<0.001; figure 3D). If we included only the CI patient groups, the relationship was even stronger (r2=0.54; p<0.0001; online supplementary figure S2). There was also a modest inverse correlation between FEV1 and mexY mRNA expression (r2=−0.39; p<0.02; data not shown).
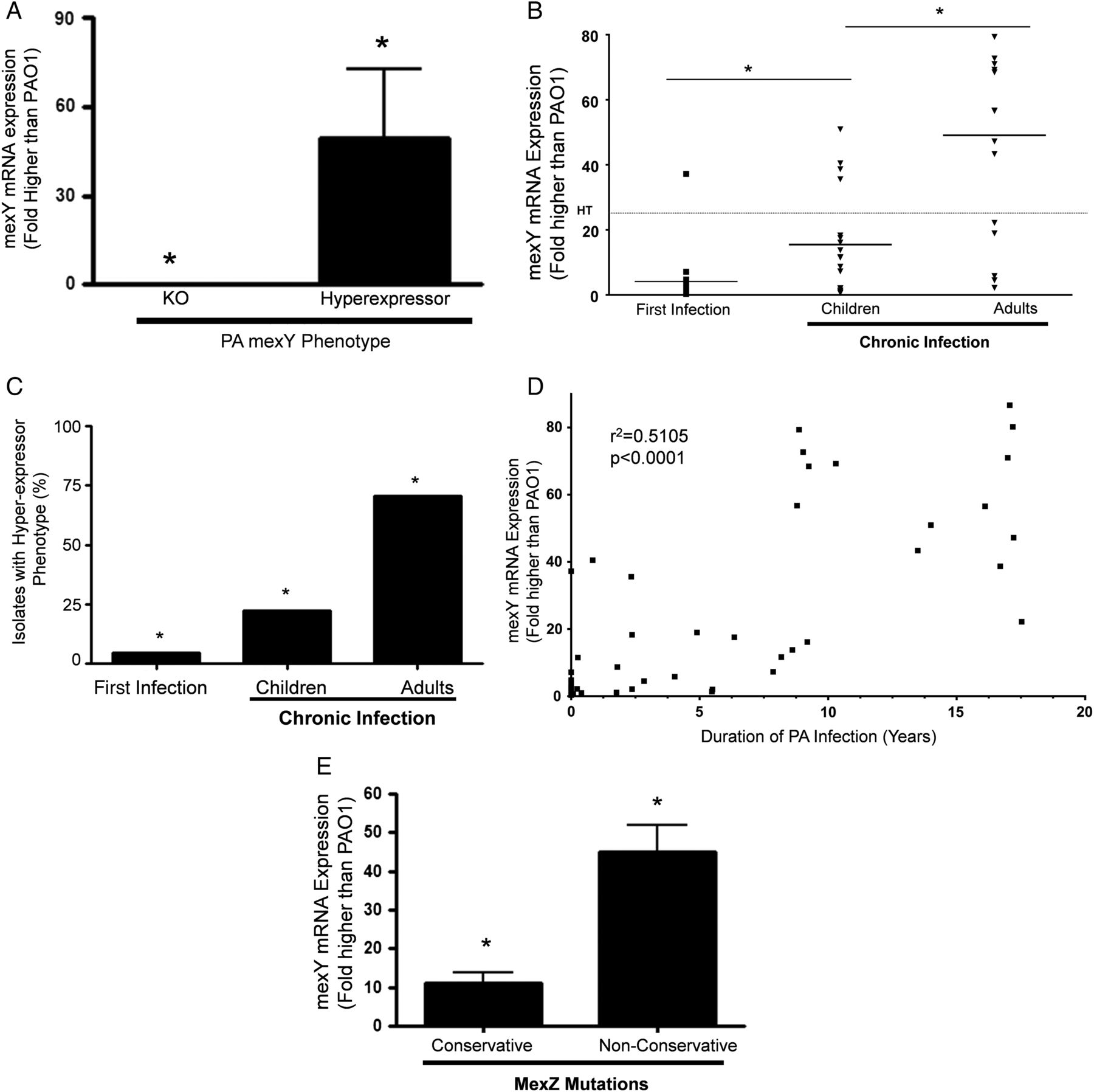
In cystic fibrosis Pseudomonas aeruginosa infection, mexY mRNA expression increases with duration of infection and is higher in isolates with non-conservative mexZ mutations. (A) MexY mRNA expression is shown in mexY knockout and mexY hyperexpressor strains (*p<0.001 for differences in mean). (B) MexY mRNA expression by P. aeruginosa isolate from each cohort as assessed by quantitative PCR is shown (*p<0.01 for differences in means between the cohorts). (C) Percentage of P. aeruginosa isolates that were hyperexpressors of mexY mRNA is shown (*p<0.01 for differences in percentage in the cohorts). (D) Relationship between duration of P. aeruginosa infection and mexY mRNA expression (r2=0.5105, p<0.0001) is shown. (E) MexY mRNA expression is shown in P. aeruginosa isolates with conservative mexZ mutations compared with non-conservative mutations (*p<0.001 for comparison of means).
We then examined the relationship between mexY mRNA expression and mexZ mutations. Median expression of mexY in P. aeruginosa isolates with non-conservative mexZ mutations was fivefold higher compared with isolates with non-synonymous conservative mexZ mutations (figure 3E). There was no difference in mexY mRNA expression in isolates with synonymous and non-synonymous conserved mutations (see online supplementary figure S3). Overall, these data are consistent with the idea that non-conserved mutations in mexZ contribute to increased mexY mRNA expression in P. aeruginosa.
Amikacin susceptibility, mexY mRNA expression and mexZ mutations
We assessed the frequency of amikacin resistance in the P. aeruginosa isolates by cohort. In the newly infected cohort, 9.5% of P. aeruginosa isolates were resistant to amikacin, compared with 22.2% of isolates from CI children and 64.7% from CI adults (p<0.001; figure 4A). We then examined the relationship between amikacin resistance, mexY mRNA expression and non-conservative mexZ mutations. As a group, the isolates that were amikacin-resistant had fivefold higher mexY mRNA expression than the isolates that were amikacin-sensitive (p<0.01; figure 4B). Further, 59% of P. aeruginosa isolates resistant to amikacin had non-conservative mexZ mutations compared with 18% of P. aeruginosa isolates that were sensitive to amikacin (p<0.01; figure 4C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pseudomonas aeruginosa amikacin resistance is higher in older patients and is associated with higher mexY mRNA expression and non-conserved mexZ mutations. (A) Percentage of P. aeruginosa isolates that are amikacin-resistant in each cohort is shown (*p<0.001 for comparison of percentages across cohorts). (B) MexY mRNA expression of P. aeruginosa isolates that were sensitive to amikacin compared with isolates resistant to amikacin is shown (*p<0.01 for comparison of means). (C) Percentage of non-conserved mexZ mutations in P. aeruginosa isolates that were amikacin-sensitive compared with those that were amikacin-resistant is shown (*p<0.01 for comparison of percentages).
Discussion
In this report, we showed that in P. aeruginosa isolates from patients with CF, amikacin resistance, mexY mRNA expression levels and mexZ mutation prevalence increase with residence time in the lung. Second, we found that mexZ mutations contribute substantially to amikacin resistance through increased mexY mRNA expression. Last, we demonstrated that mexZ mutations are generally unique to individuals, undergo positive selection and lead to changes in protein function primarily due to insertions/deletions. Overall, our results are in general agreement with previous data on a progressive increase in P. aeruginosa amikacin resistance as well as increases in MexXY expression and the prevalence of mexZ mutations in patients with CF.9 ,10 ,15
In two previous studies of patients with CF having chronic P. aeruginosa infection, the prevalence of mexZ mutations was among the highest of mutations tested, 62% and 79%, respectively.19 ,20 However, neither study reported the duration of P. aeruginosa infection or the functional consequences of mexZ mutations. Thus, the kinetics of mexZ mutations or whether they underwent positive selection cannot be ascertained from previous data. Our sequence data show that 15 of 17 non-conservative mutations were in the CTD domain or had no functional protein and only 2 of 17 were in the DBD domain. This suggests that non-conservative mutations that impact ligand–MexZ interactions with presumed secondary effects on DNA-binding are more frequent than mutations that impact DNA-binding directly. Additionally, based on the increase in both the total mexZ mutations and the ratio of non-conservative and non-synonymous mexZ mutations in cohorts with progressively greater P. aeruginosa residence time, we estimated that the prevalence of non-conservative mexZ mutations doubled every 5 years and mexZ sequence changes that alter protein function become fixed over time. This suggests that MexZ protein with altered function is integral for reaching an adaptive peak and provides a survival advantage for P. aeruginosa in the CF lung. These data support previous findings that multiple mutations may become fixed in P. aeruginosa isolates due to a positive selection pressure over the first 10 years of lung residence time.19 ,28
In addition, our data are consistent with previous studies showing in CF P. aeruginosa AG resistance increases with age and is associated with poorer outcomes.8–10 ,15 Based on the mean P. aeruginosa residence time for each cohort, we estimated that the prevalence of amikacin resistance doubled every 4–5 years of lung residence time and paralleled the increase in mexY mRNA expression and non-conservative mexZ mutations. Previous work has highlighted that AG resistance can be caused by multiple mechanisms, but the most common is by increased expression of the efflux pump MexXY.9–11 Our work provides evidence that a rising prevalence of non-conservative mexZ mutations is associated with both higher mexY mRNA expression and amikacin resistance, though it is possible to have increased MexXY expression in the absence of mexZ mutations.10 It should be noted that amikacin resistance in our assay is relevant for intravenous antibiotics only and not for inhaled antibiotics. Since inhaled antibiotics can achieve airway concentrations that are magnitudes higher than in serum, it is unknown if additional mutations are required for resistance to inhaled antibiotics.
Known factors by which increased MexXY expression may provide a fitness advantage include antibiotic exposure and oxidant stress, among others. MexXY expression can be induced in response to ribosome dysfunction, which occurs with exposure to macrolides, tetracyclines and AGs.14 ,29–31 In addition, oxidative stress can independently induce MexXY expression and AG resistance.32 Assessment of lifetime oxidative stress or collection of detailed antibiotic histories for each patient was not feasible; however, since CF lung disease progresses with age, it is well accepted that both oxidant injury and antibiotic exposure increase as patients age and FEV1 decreases.33 ,34 Though our data cannot establish causality between mexZ mutations, mexY mRNA expression and worse lung function, we did detect an inverse relationship between FEV1 and mexY mRNA expression. Further, our data are consistent with the idea that mexZ mutations may play a role in the transition from new P. aeruginosa infection to chronic infection. If true, we speculate that selection of antibiotics early in the course of CF lung disease may select for mexZ mutations and AG resistance and facilitate the transition to chronic P. aeruginosa infection.
Interestingly, there were a few P. aeruginosa isolates that were discordant with respect to mexY mRNA expression and amikacin resistance. A small percentage of P. aeruginosa isolates had high mexY mRNA expression but remained sensitive to amikacin, suggesting that there are compensatory mechanisms that can overcome high levels of MexXY.35 Conversely, some isolates had limited mexY mRNA expression but were resistant to amikacin. This can occur with mutations impacting the ribosomal-binding site of AGs or enzymatic resistance mechanisms.36
In conclusion we found that P. aeruginosa non-conservative mexZ mutations increase with longer residence time in the CF lung, likely providing a survival advantage linked to AG resistance mediated by increased MexXY expression.
References
Footnotes
Funding National Center for Advancing Translational Sciences (grant number: 8UL1TR000150); Cystic Fibrosis Foundation (grant number: Fellow Research Training Grant).
Competing interests None declared.
Ethics approval Northwestern University IRB, Lurie Children's IRB.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Airwaves