Article Text

Abstract
Background: The pathophysiology of asthma involves the action of inflammatory/allergic lipid mediators formed following membrane phospholipid hydrolysis by phospholipase A2 (PLA2). Cysteinyl leukotrienes are considered potent inducers of bronchoconstriction and airway remodelling. Ovalbumin (OVA) induced bronchoconstriction in rats is associated with increased secretory PLA2 (sPLA2) activation and cysteinyl leukotriene production, together with suppression of cytosolic PLA2 and prostaglandin E2. These processes are reversed when the animals are pretreated systemically with an extracellular cell impermeable sPLA2 inhibitor which also suppresses the early allergic reaction to OVA challenge. In this study we examine the capacity of the sPLA2 inhibitor to ameliorate inflammatory and allergic manifestations (early and late bronchoconstriction) of OVA induced allergic bronchitis in rats when the inhibitor was administered by inhalation to confine it to the airways.
Methods: Rats sensitised with OVA were treated with the sPLA2 inhibitor hyaluronic acid-linked phosphatidyl ethanolamine (HyPE). The rats were divided into four groups (n = 10 per group): (1) naïve controls (no sensitisation/no treatment); (2) positive controls (sensitisation + challenge with OVA inhalation and subcutaneous injection of 1 ml saline before each challenge; (3) sensitisation + challenge with OVA and HyPE inhalation before every challenge; and (4) sensitisation + challenge with OVA and treatment with subcutaneous dexamethasone (300 μg) before each challenge as a conventional reference. Another group received no treatment with HyPE during the sensitisation process but only before or after challenge of already sensitised rats. Pulmonary function was assessed and changes in the histology of the airways, levels of cysteinyl leukotrienes in BAL fluid, and the production of nitric oxide (No) and tumour necrosis factor α (TNFα) by BAL macrophages were determined.
Results: Inhalation of HyPE markedly suppressed OVA induced early and late asthmatic reactions as expressed by bronchoconstriction, airway remodelling (histology), cysteinyl leukotriene level in BAL fluid, and production of TNFα and NO by BAL macrophages. OVA induced bronchoconstriction in sensitised non-pretreated rats was also inhibited by inhalation of HyPE either before or after the challenge.
Conclusions: These findings confirm the pivotal role of sPLA2 in the pathophysiology of both the immediate allergic response and the inflammatory asthmatic process. Control of airway sPLA2 may be a new therapeutic approach to the treatment of asthma.
- ASM, airway smooth muscle
- CysLT, cysteinyl leukotriene
- ExPLI, extracellular sPLA2 inhibitor
- GAG, glycosaminoglycans
- HyPE, hyaluronic acid-linked phosphatidyl ethanolamine
- IL, interleukin
- LT, leukotriene
- NO, nitric oxide
- OVA, ovalbumin
- PG, prostaglandin
- PLA2, phospholipase A2
- TNFα, tumour necrosis factor α
- asthma
- experimental allergic bronchitis
- phospholipase A2
- glycosaminoglycans
Statistics from Altmetric.com
- ASM, airway smooth muscle
- CysLT, cysteinyl leukotriene
- ExPLI, extracellular sPLA2 inhibitor
- GAG, glycosaminoglycans
- HyPE, hyaluronic acid-linked phosphatidyl ethanolamine
- IL, interleukin
- LT, leukotriene
- NO, nitric oxide
- OVA, ovalbumin
- PG, prostaglandin
- PLA2, phospholipase A2
- TNFα, tumour necrosis factor α
The pathophysiology of asthma involves the action of inflammatory/allergic lipid mediators produced following hydrolysis of membrane phospholipids by phospholipase A2 (PLA2) into lyso-phospholipids and arachidonic acid. Arachidonic acid is metabolised mainly via the lipoxygenase and cyclo-oxygenase pathways, producing leukotrienes (LTs) and prostaglandins (PGs), respectively. Cysteinyl LTs (CysLTs) in particular play an important role in the pathophysiology of asthma.1 Lyso-phospholipids induce white cell activation, histamine secretion from mast cells, and airway smooth muscle (ASM) proliferation.2,3
The PLA2 family consists of intracellular and secreted enzymes;4 the intracellular enzymes include the Ca2+ independent iPLA2 which is generally considered a housekeeping enzyme, and the cytosolic cPLA2 enzyme which is specific to arachidonic acid carrying phospholipids and thus considered a major producer of arachidonic acid. Secretory PLA2 (sPLA2), secreted by activated leucocytes and other assaulted cells5,6 hydrolyse cell membrane phospholipids with no preference for fatty acid. sPLA2 also act as receptor ligands to induce production of cytokines (for example, tumour necrosis factor α (TNFα) and interleukin (IL)-6 and IL-8),7,8 mast cell survival,9 CD44 expression by eosinophils,8 and ASM cell proliferation.7 The PLA2 family is thus involved directly and indirectly in inducing inflammatory/allergic processes, especially sPLA2 which is considered the “inflammatory enzyme”.6
Although the relative contributions of cPLA2 and sPLA2 differ between cells and tissues, it is generally accepted that both types take part and often act synergistically in producing pro-inflammatory lipid mediators.10 However, the airways seem to be an exception in which different eicosanoids play opposing roles: PGE2, generally considered a strong pro-inflammatory agent, is a potent bronchodilator and can inhibit ASM proliferation,11,12 whereas CysLTs are strong bronchoconstrictors and facilitate airway remodelling.13 It has therefore been postulated that, unlike other organs, the lung is a privileged site for the beneficial actions of PGE212 and in challenged airways LTs and PGs follow different routes. Accordingly, it has been reported that, in inflammatory cells, LTs are produced from an sPLA2 linked arachidonic acid pool while PGs are produced from a cPLA2 linked pool of arachidonic acid,10,14 implying opposing roles for sPLA2 and cPLA2 in asthma.
This hypothesis was strongly supported by our recent study with an experimental allergic bronchitis model in rats15 which showed that ovalbumin (OVA) induced bronchoconstriction is associated with an increase in sPLA2 and CysLT levels and suppression of cPLA2 and PGE2 levels. Furthermore, these processes were reversed by treatment with an extracellular sPLA2 inhibitor (ExPLI), suggesting a pivotal role for sPLA2 in the pathophysiology of allergy. In this study we have tested the potential of sPLA2 inhibition in ameliorating experimental allergic bronchitis by examining the effect of ExPLI inhalation on the early and late bronchoconstriction reaction and airway inflammation and remodelling.
METHODS
Induction of experimental allergic bronchitis
Brown Norway rats were sensitised with OVA as previously described.15 Briefly, a subcutaneous injection of OVA with aluminium hydroxide and an intraperitoneal injection of Bordetella pertussis killed organisms were administered on day 0. Two weeks later the rats were placed unrestrained in a 20 litre box connected to an ultrasonic nebuliser (LS 230 System, Villeneuve sur Lot, France) and subjected to repeated bronchial allergen challenge by inhalation of OVA (1 mg/ml saline) for 5 minutes every other day to day 45.
Treatments
The ExPLI used was hyaluronic acid-linked phosphatidyl ethanolamine (HyPE) which was synthesised in the laboratory of S Yedgar16 by conjugating hyaluronic acid (HA, Gideon Richter, Budapest, Hungary) with dipalmitoyl phosphatidyl ethanolamine (PE, Sigma, St Louis, MI) to form an amide bond between the PE amino group and the HA carboxylic group, and was characterised by NMR spectroscopy.16,17
Two treatment modes were applied:
-
HyPE inhalation during the induction of allergic bronchitis to examine the effect of ExPLI on sensitisation (prevention). Four treatment groups (each of 10 rats) were used:
naïve control (−/−): no sensitisation/no treatment;
positive control (OVA/OVA): sensitisation + challenge with OVA inhalation and subcutaneous injection of 1 ml saline before each challenge;
sensitisation + challenge with OVA and HyPE inhalation before every challenge (OVA/HyPE);
sensitisation + challenge with OVA and treatment with subcutaneous dexamethasone (300 μg) before each challenge (OVA/Dx) as a conventional reference.
-
No treatment with HyPE during the sensitisation process but only before or after challenge of already sensitised rats.
Five ml HyPE solution (1 mg/ml saline) was sprayed into the 20 l cage, thus diluting the HyPE to 0.25 μg/ml aerosol, and the rats inhaled the aerosol for 5 minutes. The respiratory rate of the rats was 120 breaths/min with a mean tidal volume of 1 ml, giving a ventilation rate of 120 ml/min. If all the inhaled HyPE was absorbed in 5 minutes (600 ml) the maximal HyPE absorbed would be 150 μg (3 nmol) in each treatment. Each group was treated and challenged every other day from day 14 until day 45.
Assessment of pulmonary function
Unrestrained conscious rats were placed in a whole body plethysmograph (Buxco Electronics, Troy, NY, USA) to measure flow derived pulmonary function (Penh) as described elsewhere.15,18 Bronchoconstriction was assessed on day 20 by measuring Penh before and 5 minutes and 8 hours after OVA challenge, corresponding to the early and late asthmatic reaction, respectively.
Histological examination
On day 45 the rats were anaesthetised with an intraperitoneal injection of sodium pentobarbital, sacrificed by bleeding through the abdominal aorta, tracheotomised and cannulated through the trachea. Bronchoalveolar lavage (BAL) fluid was collected by repeated washing of the lungs with 5 ml saline to a total of 50 ml. The lungs were removed, inflated, fixed with 4% formaldehyde under a pressure of 20 cm H2O, and embedded in paraffin. The fixed lungs were sliced longitudinally and embedded in paraffin. Histological sections 3 µm thick were cut and stained with (1) haematoxylin and eosin for assessment of interstitial and peribronchial inflammation and of ASM thickening; (2) Tri-chrome for assessment of subepithelial fibrosis (basal membrane thickness); and (3) Periodic acid Schiff (PAS) for examination of epithelial cell mucus metaplasia.
Histological morphometry was performed using the computer program ImageJ (NIH, Bethesda, MD, USA) on three randomly selected slides from each lung. Quantification of peribronchial cellular infiltrate in airway tissue was determined by counting the numbers of inflammatory cells in the region 50 μm beneath the epithelium of the airway in haematoxylin and eosin stained sections and expressed as the number of cells per mm airway basal lamina length which was measured by tracing the basal lamina in calibrated digital images. ASM and basal membrane thickening were determined as previously described.18 Briefly, the area of ASM cells or the basal membrane thickness was normalised to the square of the basement membrane length (Lbm2, µm2) to correct for differences in airway size. Mucus metaplasia was determined by the percentage of PAS positive cells in the total airway epithelial cells. Only large (>2000 µm Lbm) and medium size airways (1000−2000 µm Lbm) were selected as it has been found that the most significant pathological changes occur in these airways with less significant changes in the small airways.19 Airways with a ratio of maximum to minimum internal diameter of ⩾2 were considered to be cut tangentially and were not measured. The internal diameters were measured from the basement membrane on one side to the basement membrane on the opposite side of the airway.
Biochemical markers
CysLTs (LTC4, LTD4 and LTE4) were determined by BAL enzyme immunoassay using a commercial kit (Amersham, UK).15
To measure production of nitric oxide (NO) and tumour necrosis factor α (TNFα) by macrophages, BAL macrophages were isolated and cultivated in DMEM + 10% serum for 2 hours, after which non-adherent cells were removed. The adherent cells were recultivated in DMEM + 10% serum (106 cells/well) for 48 hours. The culture medium was then collected for determination of NO using the Gries method for measuring the corresponding nitrite, and TNFα using a radioimmunoassay kit (Amersham, UK).
Analysis of data
Statistical analysis was performed using statistical software (GB-STAT, Dynamic Microsystem, Silver Spring, MD, USA). Analysis of variance (ANOVA) was used to assess significant differences between treatment groups. The Tukey test was used to compare each of the treatment groups. A p value of <0.05 was considered significant.
RESULTS
Effect of HyPE inhalation on respiratory function in rats with OVA induced experimental allergic bronchitis
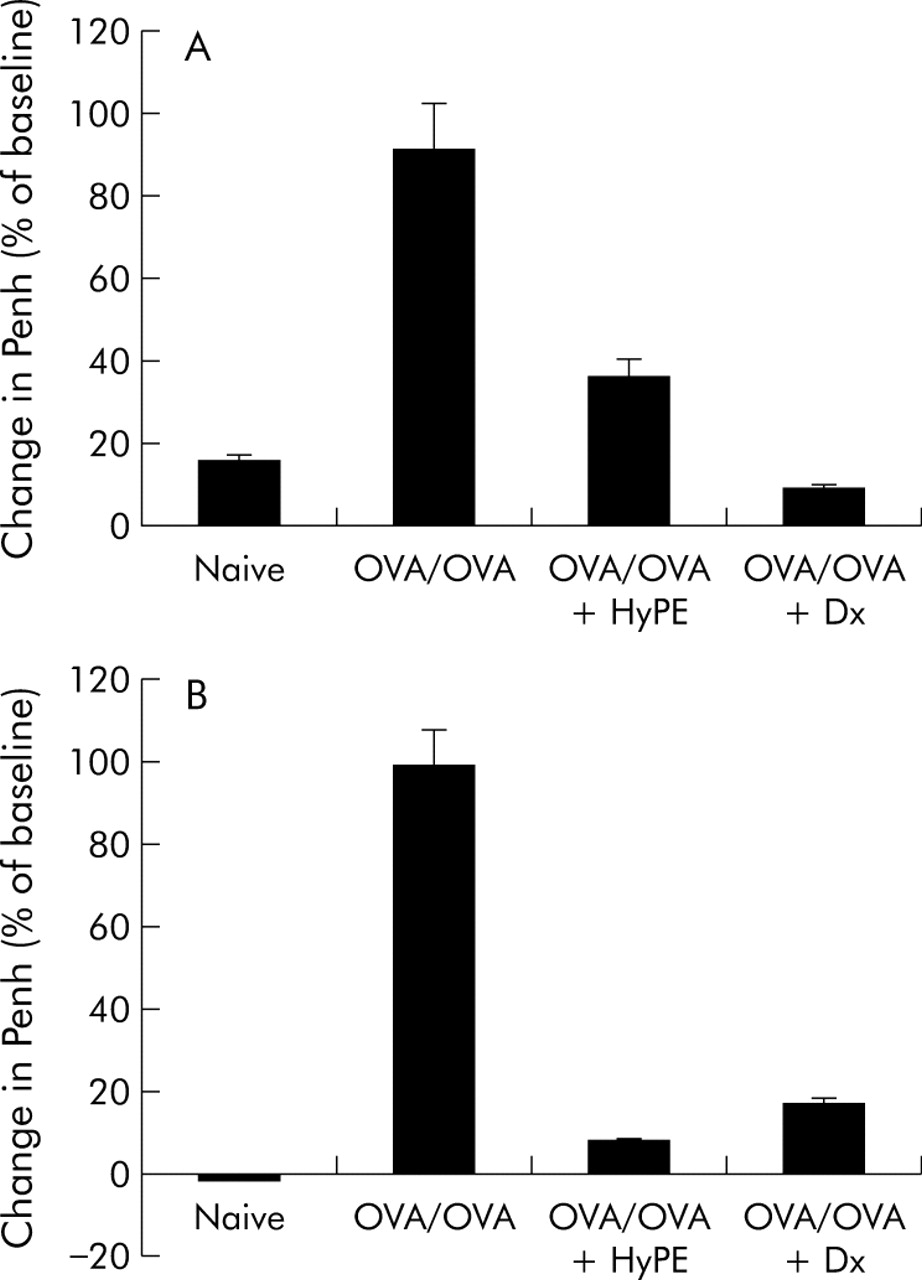
To examine the effect of ExPLI on sensitisation of rats by OVA, the rats inhaled HyPE before every challenge as described above. In parallel, another group was given a subcutaneous injection of dexamethasone (Dx). As shown in fig 1, inhalation of HyPE was effective in reducing bronchoconstriction in both the early and late reactions.
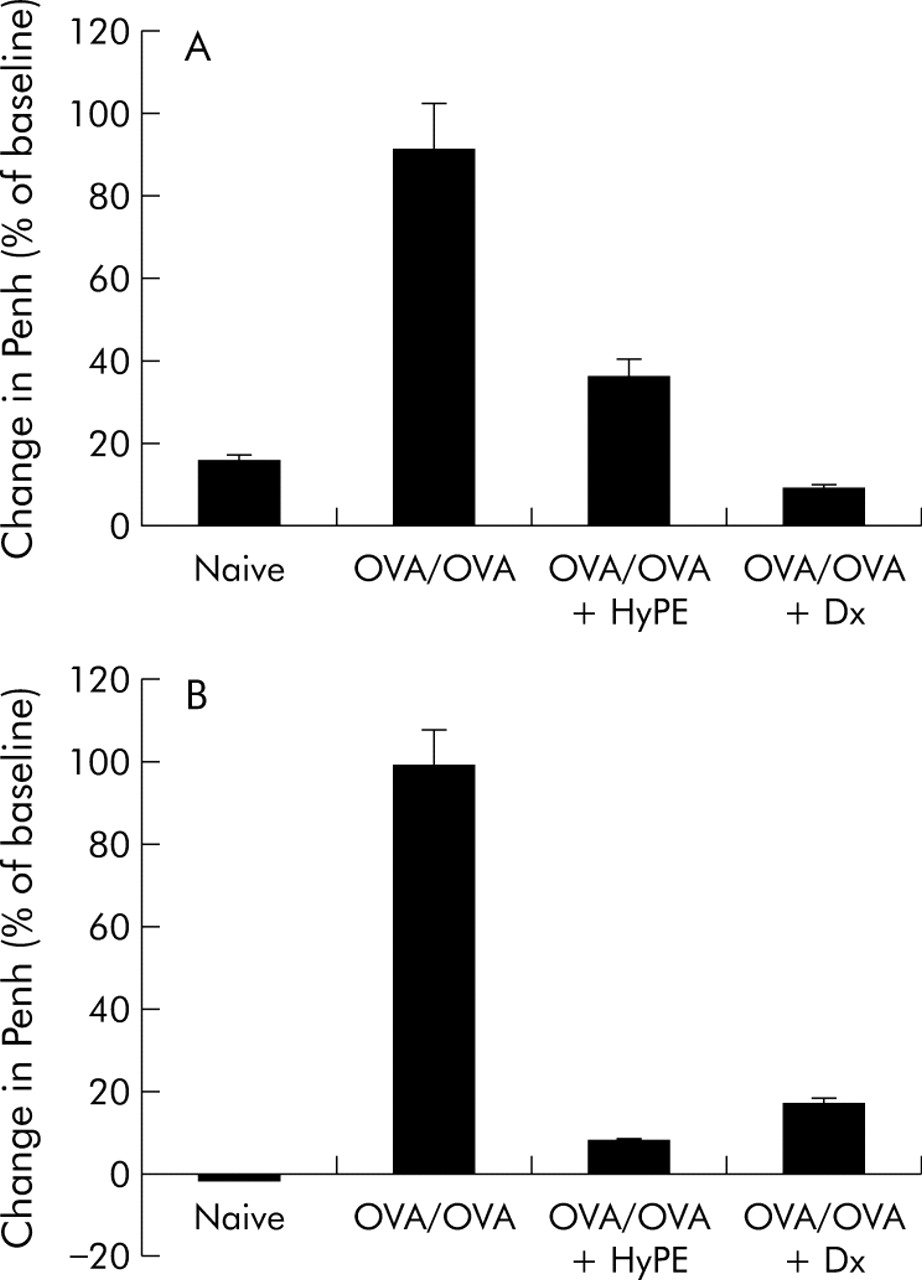
Effect of HyPE inhalation on (A) early and (B) late bronchoconstrictive reactions induced by ovalbumin (OVA) inhalation. Bronchoconstriction, expressed as the percentage change in Penh,16 was induced in OVA sensitised rats by inhalation of OVA and measured before, 5 minutes after, and 8 hours after allergen challenge. Data shown are mean (SE) values for 10 rats. p<0.01 between OVA/OVA and the other groups for both early and late reactions.
To examine the ability of inhaled HyPE to counteract OVA induced bronchoconstriction in untreated allergic reaction, rats sensitised with OVA but not pretreated with HyPE inhaled HyPE only once, either before or after challenge with OVA. Figure 2 shows that HyPE decreased OVA induced bronchoconstriction when inhaled either before or after the challenge.

Immediate effect of HyPE inhalation on ovalbumin (OVA) induced bronchoconstriction in OVA sensitised rats. (A) Prevention of bronchoconstriction: OVA sensitised rats (not treated with HyPE during sensitisation) were subjected to Penh determination at baseline, 5 minutes after inhalation of HyPE (diamonds) or saline (squares), and 5 minutes after OVA inhalation. (B) Relief of bronchoconstriction: OVA sensitised rats (not treated with HyPE during sensitisation) were subjected to Penh determination at baseline, 5 minutes after inhalation of OVA, and 5 minutes after inhalation of HyPE (diamonds) or saline (squares). Data points are mean (SE) for five rats. In (A) p<0.05 between HyPE and saline treated rats and OVA+HyPE treated rats. In (B) p<0.01 between HyPE and saline treated rats.
Prevention of airway remodelling by HyPE inhalation
To examine the effect of HyPE inhalation on allergic bronchial inflammation, at the end of the experiment (30 days after treatment and repeated challenge) the rats were sacrificed and tracheotomised, BAL fluid was collected, and lung tissues were processed for histological examination.
As shown in fig 3, peribronchiolar and perivascular cell infiltrates were composed principally of eosinophils and lymphocytes. The eosinophils were distributed in a circumferential manner around the bronchioles and vessels while the lymphocytes were either similarly distributed or arranged in primary follicles (lymphoid follicles without germinal centres). Reduced inflammatory cell infiltration and airway wall thickening was seen in HyPE treated rats. Table 1 shows that inhalation of HyPE reduced peribronchial cellularity, mucus metaplasia of respiratory epithelial cells, ASM thickening, and basal membrane thickening essentially to the level of the naïve rats.
Effect of HyPE on remodelling of OVA sensitised rat airways: histological morphometry
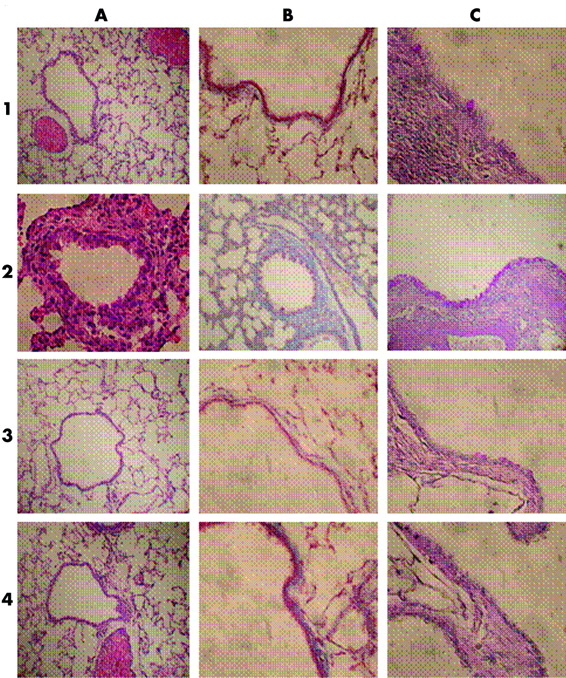
Effect of HyPE inhalation on structural changes in airways (airway remodelling) of OVA sensitised rats. Rats were subjected to OVA inhalation every other day for 30 days. Rats treated with HyPE inhaled HyPE aerosol for 5 minutes before each allergen inhalation. The rats were sacrificed on day 45. (A) Haematoxylin-eosin stain for detection of inflammatory cell infiltration and changes in airway smooth muscle cell (ASM) thickness. (B) Mason-Trichrom stain of connective tissue (collagen) for detection of changes in basal membrane thickness. (C) Periodic Acid Schiff (PAS) stain for detection of mucus metaplasia of respiratory epithelial cells. 1, 2, 3 and 4 indicate tissues of naïve, OVA/OVA, HyPE treated, and dexamethasone treated rats, respectively.
Effect of HyPE inhalation on biochemical markers
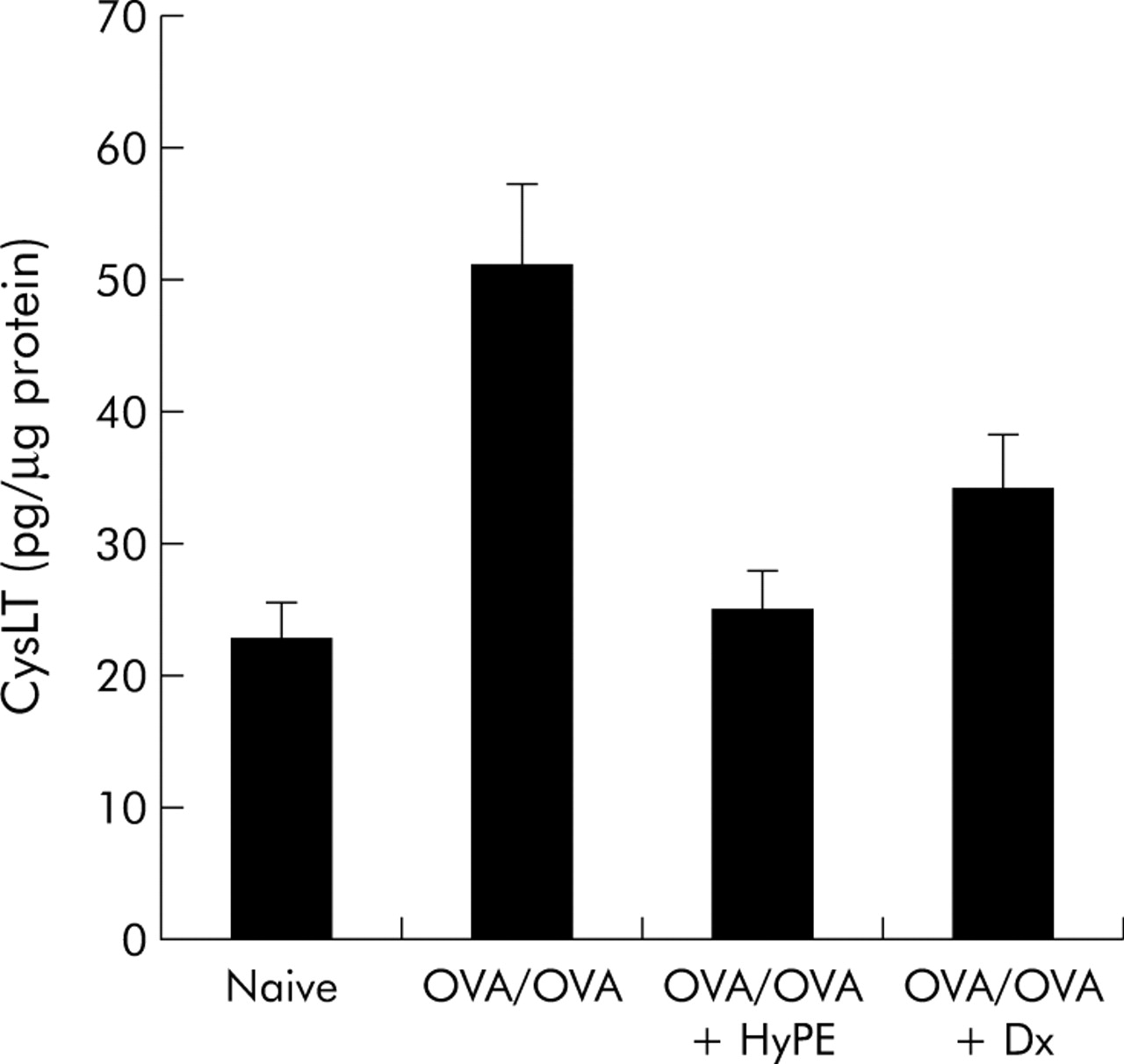
Among the lipid mediators involved in asthma pathophysiology, CysLTs are considered key mediators of bronchoconstriction and the inhibition of their action is currently the target of several drugs (montelukast, zileuton) used for the treatment of asthma.20 As the PLA2 inhibitor controls lipid mediator production, it was of particular interest to determine its effect on CysLT production in OVA sensitised rats. CysLT levels in the BAL fluid of the different groups were determined by ELISA. As shown in fig 4, CysLT production was markedly enhanced in sensitised rats but was reduced to the normal range by inhalation of HyPE.

Effect of HyPE inhalation on cysteinyl leukotriene (LTC4, LTD4 and LTE4) levels in BAL fluid of ovalbumin (OVA) sensitised rats. BAL fluid was collected upon sacrifice and CysLT levels were determined by enzyme immunoassay. Data are mean (SE) values for 10 rats. A significant difference was seen between OVA/OVA and HyPE treated rats (p<0.01). No significant differences were seen between HyPE treated and naïve or dexamethasone (Dx) treated rats.
Nitric oxide production is a characteristic marker of inflammation in asthma which correlates with disease severity, although it might have dual effects on mast cells.21 Its production has been reported to be linked to PLA2 produced lipid mediators22 and was found to be inhibited by ExPLIs in lipopolysaccharide stimulated cells23 and animals.24 In line with these findings, we determined the effect of ExPLI on NO production by cultured macrophages collected from BAL fluid of the different groups following their in vivo treatment without further treatment of the cultured cells. Figure 5 shows that NO production was considerably enhanced in BAL macrophages of sensitised rats (OVA/OVA) compared with naïve (−/−) rats, but NO production by macrophages from sensitised rats treated with HyPE inhalation was below the level of the naïve rats.

Effect of HyPE inhalation on nitric oxide (NO) production by macrophages collected from BAL fluid of ovalbumin (OVA) sensitised rats. Macrophages collected from BAL fluid of the different groups were cultured without further treatment with HyPE or dexamethasone (Dx) and NO production was determined by the corresponding nitrate level in the culture medium. Data are mean (SE) values for 10 rats. The NO level was reduced compared with OVA/OVA and naïve rats by both HyPE (p<0.001 and p<0.001, respectively) and Dx (p<0.001 and p<0.001, respectively).
A prominent characteristic marker of inflammatory conditions is TNFα which is at the top of other pro-inflammatory cytokine cascade. The increase in its level correlates with the severity of inflammation in asthma, and its inhibition has been proposed for the treatment of severe asthma.25 In previous studies the ExPLIs were found effective in suppressing endotoxin stimulated production of TNFα and other cytokines in animals and cell cultures.24,26 Similarly, we found that TNFα production by macrophages from BAL fluid of sensitised rats was markedly enhanced, but inhalation of HyPE reduced it to approximately the level of the naïve rats (fig 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of HyPE inhalation on tumour necrosis factor α (TNFα) production by macrophages collected from BAL fluid of ovalbumin (OVA) sensitised rats. Macrophages collected from BAL fluid of the different groups were cultured without further treatment with HyPE or dexamethasone (Dx) and secretion of TNFα into the culture medium was determined. Data are mean (SE) values for 10 rats. p<0.001 for difference between OVA/OVA and HyPE treated rats. There was no significant difference between HyPE treated, naïve, and Dx treated rats.
DISCUSSION
Inhibition of PLA2 activity has been considered a promising therapeutic strategy in the treatment of inflammatory/allergic diseases as it can suppress the production of several classes of pathogenic lipid mediators (PGs, LTs, platelet aggregating factor, lysophospholipids). A number of PLA2 inhibitors have been proposed for this purpose.27 However, PLA2 inhibitors that enter the cell might interfere with the vital phospholipase metabolism and might impair the cell viability. In addition, as previously discussed,6 in inflammatory conditions a number of sPLAs are involved in cell lysis and lipid mediator production, and inhibiting only one or part of them has limited or no efficacy. Therefore, as has long been proposed,28 there is a clear advantage to cell impermeable PLA2 inhibitors that protect the cell membrane from the action of sPLA2s but are not internalised.6 These requirements are provided by the ExPLIs which are composed of a lipidic PLA2 inhibitor conjugated with a polymeric carrier.17,29 While the lipidic component is incorporated into the cell membrane, its internalisation is prevented by the polymeric carrier. These kind of lipid conjugates have been shown to protect membranes from diverse types of sPLA2s29,30 and ameliorate different sPLA2 related inflammatory conditions including endotoxin induced sepsis in rats,24 allergic experimental encephalomyelitis in rats and mice,23 and TNBS induced colitis in rats.31
We have previously shown that sensitisation of rats with OVA is associated with an increase in sPLA2 and CysLT production together with suppression of cPLA2 and PGE2 production. These processes were reversed by systemic (subcutaneous) treatment with the cell impermeable PLA2 inhibitor HyPE which also suppressed the early response (bronchoconstriction) to challenge with OVA.15 The present study shows that effective treatment of asthma can be obtained by application aimed at confining the treatment to the airway system. Inhalation of HyPE by asthmatic rats suppressed OVA induced sensitisation and airway inflammation (fig 1, table 1) while at the same time blocking CysLT production (fig 4), and also reversed OVA induced bronchoconstriction in already sensitised rats (fig 2). Together, the present and preceding studies provide strong evidence for the pivotal role of sPLA2 in the pathophysiology of both the immediate allergic response and the inflammatory process.
Previous studies have pointed out the involvement of PLA2 in NO production, showing that exogenous PLA2 induces iNO synthase (iNOS) expression and NO generation by alveolar macrophages, a phenomenon that contributes to lung injury.22 In accordance with this we have found that treatment of rat glial cells with ExPLIs inhibits LPS induced production of NO concomitantly with PGE2 production and PLA2 secretion as measured by its lipolytic activity.23 In a study of the effect of ExPLI on sepsis in rats, we found that treatment with HyPE suppressed expression of iNOS and sPLA2-IIA mRNA in lung and kidney of rats with endotoxin induced sepsis.24 The present study provides further evidence for the relationship between NO production and PLA2 activity, showing that treatment with the extracellular PLA2 inhibitor suppressed the capacity of BAL macrophages to produce NO (fig 5).
The marked reduction in the ability of BAL macrophages to produce TNFα following in vivo treatment of asthmatic rats with HyPE, without further treatment of the cultured cells (fig 6), may be attributed to PLA2 activity. However, the dependence of TNFα production on PLA2 enzymatic activity is unclear. In a previous study from our laboratory it was found that ExPLIs inhibited lipopolysccharide induced production of different cytokines in cultured endothelial cells,26 which would suggest a relation to PLA2 activity. However, in that study we could not prove a dependence of cytokine production on PLA2 lipolytic activity. On the other hand, treatment with ExPLIs inhibited lipopolysaccharide induced activation of NFκ-B which mediates TNFα production in cultured endothelia cells26 as well as TNFα production in septic rats.24 As discussed above, sPLA2 enzymes can induce cytokine production via a receptor mediated process which is independent of their enzymatic activity.7 It is thus possible that the effect of ExPLIs on cytokine production is due to suppression of PLA2 action as a receptor ligand rather than its enzymatic activity.
The lipolysis independent effects of the ExPLIs may be attributed to the fact that they are membrane anchored glycosaminoglycans (GAGs) or GAG-like molecules. Cell surface GAGs are known to protect the cell from exogenous damaging agents such as endotoxins, reactive oxygen species, degradation enzymes (for example, heparanase), and cytokines.32,33 A number of studies have shown that the action of sPLA2 on the cell membrane is inhibited by cell surface GAGs such as heparin34 and hyaluronic acid.29,35 In a study of the synergistic action of hydrogen peroxide and sPLA2 we have previously observed that only after the cell surface GAGs are degraded by hydrogen peroxide is the cell membrane rendered accessible to lysis by exogenous sPLA2.32 Although the relationship between GAG function and the action of inflammatory mediators is not unequivocally clear, it is well documented that GAG stripping exposes cells and tissues to damaging agents. Accordingly, it has been proposed that enrichment of cell surface GAGs would assist in protecting the cell from infection and inflammatory/allergic assaults.36 These requirements are fulfilled by the ExPLIs, which are composed of PLA2 inhibiting molecules such as N-derivatised phosphatidyl ethanolamine to polymeric carriers (for example, hyaluronic acid or chondroitin sulfates). Because of their structure, these lipid conjugates exhibit a multiple effect in cell protection. The lipid moiety, which incorporates into the cell membrane, suppresses membrane phospholipase hydrolysis by endogenous sPLA2 and the subsequent production of lipid mediators23,29,30,31 while, at the same time, the polymeric carrier mimics the cell surface GAG in protecting cells from exogenous sPLA2 and other inflammatory agents such as reactive oxygen.32,35 The soundness of this concept has been demonstrated in several in vivo studies, as cited above.23,24,31 Of the different GAGs, hyaluronic acid is of particular relevance to the present study since it specifically binds to CD44 on lymphocytes and eosinophils and activates them. Accordingly, binding of hyaluronic acid to lymphocyte CD44 prevents their rolling and subsequent extravasation to inflammatory sites.37 This makes HyPE-like lipid conjugates particularly suitable for the treatment of asthma, and this indeed is shown by the findings of the present study.
In conclusion, the results of this study suggest a novel therapeutic approach to the treatment of allergic/inflammatory diseases and present the lipid conjugates—which concomitantly control PLA2 action and mimic cell surface GAG function—as a prototype of potential multifunctional non-steroidal inhaled drugs for the comprehensive treatment of asthma.
REFERENCES
Footnotes
-
Published Online First 30 June 2005
-
This study was supported by a grant to S Yedgar from the Walter and Greta Stiel Chair and a grant to D Shoseyov and S Yedgar from the Israel Ministry of Health.
-
Competing interests: none declared.