Article Text

Abstract
Background: A study was undertaken to investigate the pathogenesis of pulmonary involvement in human T lymphotropic virus type I (HTLV-I) carriers.
Methods: The bronchoalveolar lavage (BAL) cell profile of 30 HTLV-I carriers (15 asymptomatic HTLV-I carriers (AHCs) and 15 symptomatic HTLV-I carriers (SHCs)) with chronic inflammatory diseases of respiratory tract and eight patients with HTLV-I associated myelopathy/tropical spastic paraparesis (HAM/TSP) was investigated. The HTLV-I proviral deoxyribonucleic acid (DNA) load in peripheral blood mononuclear cells (PBMCs) and BAL fluid from HTLV-I carriers was estimated using the quantitative polymerase chain reaction method and the correlation between the lymphocyte number in BAL fluid and the HTLV-I proviral DNA load in PBMCs and BAL fluid was examined.
Results: The percentage of lymphocytes in BAL fluid was increased (>18%) in 11 of 30 HTLV-I carriers although there was no significant difference compared with control subjects. In HTLV-I carriers the lymphocyte number in BAL fluid correlated well with the copy number of HTLV-I proviral DNA in PBMCs. In addition, the copy number of HTLV-I proviral DNA in BAL fluid correlated well with the number of lymphocytes (both CD4+ and CD8+ cells) in BAL fluid.
Conclusions: These findings suggest that pulmonary lymphocytosis can occur in a subset of HTLV-I carriers without HAM/TSP and that the increased HTLV-I proviral DNA load may be implicated in the pathogenesis of pulmonary involvement in HTLV-I carriers.
- AHC, asymptomatic HTLV-I carrier
- BAL, bronchoalveolar lavage
- HAM/TSP, HTLV-I associated myelopathy/tropical spastic paraparesis
- HTLV-I, human T lymphotropic virus type I
- PBMC, peripheral blood mononuclear cell
- PCR, polymerase chain reaction
- SHC, symptomatic HTLV-I carrier
- bronchoalveolar lymphocytosis
- human T lymphotropic virus type I (HTLV-I)
Statistics from Altmetric.com
- AHC, asymptomatic HTLV-I carrier
- BAL, bronchoalveolar lavage
- HAM/TSP, HTLV-I associated myelopathy/tropical spastic paraparesis
- HTLV-I, human T lymphotropic virus type I
- PBMC, peripheral blood mononuclear cell
- PCR, polymerase chain reaction
- SHC, symptomatic HTLV-I carrier
Human T lymphotropic virus type I (HTLV-I) is a type C retrovirus that is aetiologically associated with adult T cell leukaemia1,2 and with HTLV-I associated myelopathy/tropical spastic paraparesis (HAM/TSP).3,4 In addition to these diseases, a number of inflammatory disorders have also been described in association with HTLV-I including HTLV-I uveitis,5 arthropathy,6 and Sjögren’s syndrome.7 Pulmonary involvement is also associated with HTLV-I—for example, in patients with HAM/TSP and HTLV-I uveitis pulmonary involvement may be characterised by bronchoalveolar lymphocytosis.8,9,10,11 Furthermore, a few preliminary studies have shown that similar pulmonary involvement is observed in HTLV-I carriers who have not developed HAM/TSP or HTLV-I uveitis.10,12
Many kinds of immunological abnormalities and an increased HTLV-I proviral deoxyribonucleic acid (DNA) load in peripheral blood, cerebrospinal fluid, and bronchoalveolar lavage (BAL) fluid from patients with HAM/TSP13,14 and HTLV-I uveitis11 have been reported, suggesting that immunological mechanisms related to an increased amount of HTLV-I proviral DNA may be implicated in the pathogenesis of these diseases. However, despite advances in elucidating the pathophysiology of these diseases, much of the information on the pathogenesis is confined to HAM/TSP and HTLV-I uveitis. There is little information available regarding pulmonary involvement and pathophysiology in HTLV-I carriers who have not developed HAM/TSP or HTLV-I uveitis.
To examine the incidence and pathogenesis of pulmonary lymphocytosis in HTLV-I carriers, we have analysed BAL cell profiles in HTLV-I carriers including asymptomatic HTLV-I carriers (AHCs). We also estimated the HTLV-I proviral DNA load in peripheral blood mononuclear cells (PBMCs) and BAL cells from HTLV-I carriers by the quantitative polymerase chain reaction (PCR) method and examined the correlation between the HTLV-I proviral DNA load and pulmonary lymphocytosis.
METHODS
This study was reviewed and approved by the Kagoshima University Faculty of Medicine Committee on Human Research.
Study subjects
The study subjects consisted of 30 HTLV-I carriers and eight patients with HAM/TSP consecutively presenting to our department between 1989 and 2000. The 30 HTLV-I carriers consisted of 15 AHCs (three men and 12 women) and 15 symptomatic HTLV-I carriers (SHCs; five men and 10 women) as shown in table 1. There were no significant differences in age between each of the groups and the control subjects. All subjects were seronegative for human immunodeficiency virus (HIV) 1.
Clinical background and bronchoalveolar lavage (BAL) findings of HTLV-I carriers and patients with HAM/TSP
To assess the cellular characteristics of BAL fluid in AHCs, the serum anti-HTLV-I antibody was checked in individuals consulting our department for an annual chest radiograph. The anti-HTLV-I antibody was measured by the gelatin particle agglutination method (Fujirebio, Tokyo, Japan). After obtaining informed consent, further examinations including fibreoptic bronchoscopy were performed on HTLV-I seropositive individuals along with careful history taking including occupational history. Individuals who worked in environments known to cause allergic lung diseases were excluded. After the diagnostic procedure, 15 AHCs were recruited to the study (table 1); the chest radiographic findings were normal in 13 individuals and two had minimal inactive tuberous lesions.
The 15 SHCs (subjects 16–30) were recruited from the outpatient clinic of our department for chronic inflammatory diseases of the respiratory tract (three sinobronchial syndrome, three bronchiectasis, two middle lobe syndrome, and two chronic bronchitis); two had an inactive tuberculous lesion and three (subjects 16–18) complained of a slight cough for 1–3 months during the study. Eight patients with HAM/TSP (three men and five women) had been diagnosed according to the criteria proposed by Osame et al.15 Three HTLV-I carriers (subjects 9, 18 and 25) were current smokers, three HTLV-I carriers (subjects 2, 8 and 23) were ex-smokers with intervals ranging from 3 months to 5 years since smoking cessation, and the others had never smoked.
White blood cell counts in peripheral blood were within the normal range in all subjects. The serum anti-HTLV-I antibody titre ranged from 128 to 16384× in HTLV-I carriers and from 512 to 32768× in patients with HAM/TSP. The median (range) % vital capacity of AHCs, SHCs, and patients with HAM/TSP was 104.9% (77.2–131.8), 85.8% (75.9–131.3), and 96.7% (76.7–108.0), respectively, and the % forced expiratory volume in 1 second in the three groups was 82.2% (79.3–91.7), 70.7% (51.6–86.2), and 79.7% (70.0–88.3%), respectively. There were no significant differences in pulmonary function between the three groups.
Nine healthy individuals (three men and six women) of median age 53 years, all non-smokers, who were seronegative for HTLV-I acted as controls. They included four healthy volunteers and five healthy individuals undergoing an annual chest radiographic examination. These latter five healthy individuals were finally diagnosed as having a small solitary lung nodule without signs of pulmonary disease (n = 3) or minimal inactive tuberculous lesions (n = 2). BAL was performed to diagnose the small lung nodule and the cellular characteristics of the BAL fluid from these subjects served as control values for this study. The chest radiographic findings of the five healthy volunteers were normal.
Bronchoalveolar lavage
Informed consent was obtained from all individuals before they underwent BAL which was performed before interventions including corticosteroid administration. Under local anaesthesia with 2% lidocaine, a fibreoptic bronchoscope was placed in the subsegment of the right middle lobe or lingua and 160 ml sterile saline was infused in four aliquots through the bronchoscope and aspirated by gentle hand suction. The lavage fluid obtained was passed through two sheets of sterile gauze and a 10 ml aliquot was centrifuged at 400g for 10 minutes, stained with Wright-Giemsa stain, and the cell differentials were determined (at least 500 cells were counted). The lavage fluid was then washed twice and the total number of cells counted.
An aliquot of BAL cells was used for identification of T lymphocyte subsets. The cells were washed twice with phosphate buffered saline and incubated with an optimal concentration of fluorescence conjugated monoclonal antibodies (OKT4 (CD4), OKT8 (CD8); Ortho Diagnostics, Raritan, NJ, USA). The cells were then analysed for surface fluorescence using flow cytometry (FCMID, Nihon Bunko, Tokyo, Japan).
Isolation of peripheral blood mononuclear cells (PBMCs)
The PBMCs were isolated from 30 ml heparinised peripheral blood by Ficoll-Hypaque density gradient centrifugation (Pharmatica, Uppsala, Sweden). Blood samples were obtained before performing BAL in all subjects. These samples were stored in liquid nitrogen until use.
Quantitative PCR of PBMCs and BAL cells
Quantitative PCR assay was performed as previously described.16 The amount of HTLV-I proviral DNA was calculated using the following formula: copy number of HTLV-I (pX) per 104 PBMCs and per 104 BAL cells = [(copy number of pX)/(copy number of β-actin/2)] × 104
Detection of HTLV-I proviral DNA from BAL cells by PCR
To examine the presence of HTLV-I proviral DNA in BAL cells, isolated BAL cells from 10 initial AHCs (nos 1–9 and 11) were analysed. As negative and positive controls, BAL cells and HTLV-I infected TCL-Kan cells17 from HTLV-I seronegative controls were also assayed. 1 μg of DNA extracted from BAL cells was used for the PCR. The amplification reaction of PCR was performed for 35 cycles and consisted of denaturation at 94°C for 1 minute, annealing at 65°C for 2 minutes, and primer extension at 72°C for 3 minutes. 10 μl of the amplified products was blotted onto a nylon membrane and hybridised with a biotin labelled probe for HTLV-I pX. After incubation with streptavidin-alkaline phosphate conjugate, disodium 3-(4-methoxyspirol [1,2-dioxetane-3-2′-tricyclo-[3.3.1.1.3.7] decan]-4-yl)phenyl phosphate (AMPPD)18 (Southern Light Kit, Troix) was added to the blot to a final concentration of 0.25 mM and the immersed blot was then slowly agitated for 5 minutes. Finally, the chemiluminescent signal was detected by exposing the radiographic film.
The location and sequences of the primers and probe are summarised in table 2.
Oligonucleotides for PCR detection of HTLV-I proviral DNA
Statistical analysis
All values are shown as median (range). Statistical analysis was performed using the Mann-Whitney U test and Spearman rank correlation. p values of <0.05 were considered significant.
RESULTS
Cellular characteristics and T lymphocyte subset of BAL fluid
The recovery ratios of BAL fluid in HTLV-I carriers, AHCs, SHCs, and patients with HAM/TSP did not differ significantly from those of control subjects (data not shown). As shown in table 3, the BAL fluid cell count was increased in HTLV-I carriers, AHCs, SHCs, and patients with HAM/TSP compared with control subjects. The differential cell count showed an increased percentage of lymphocytes and a decreased percentage of macrophages in BAL fluid from patients with HAM/TSP compared with control subjects. The percentage of lymphocytes in BAL fluid was increased (>18%) in four AHCs and seven SHCs but there was no significant difference from that in control subjects.
Bronchoalveolar lavage findings in HTLV-I carriers and HAM/TSP patients
Analysis of T lymphocyte subsets in BAL fluid showed an increased percentage of CD8+ cells in patients with HAM/TSP and an increased ratio of CD4/CD8 in AHCs.
HTLV-I proviral DNA in PBMCs and correlation with lymphocytes in BAL fluid
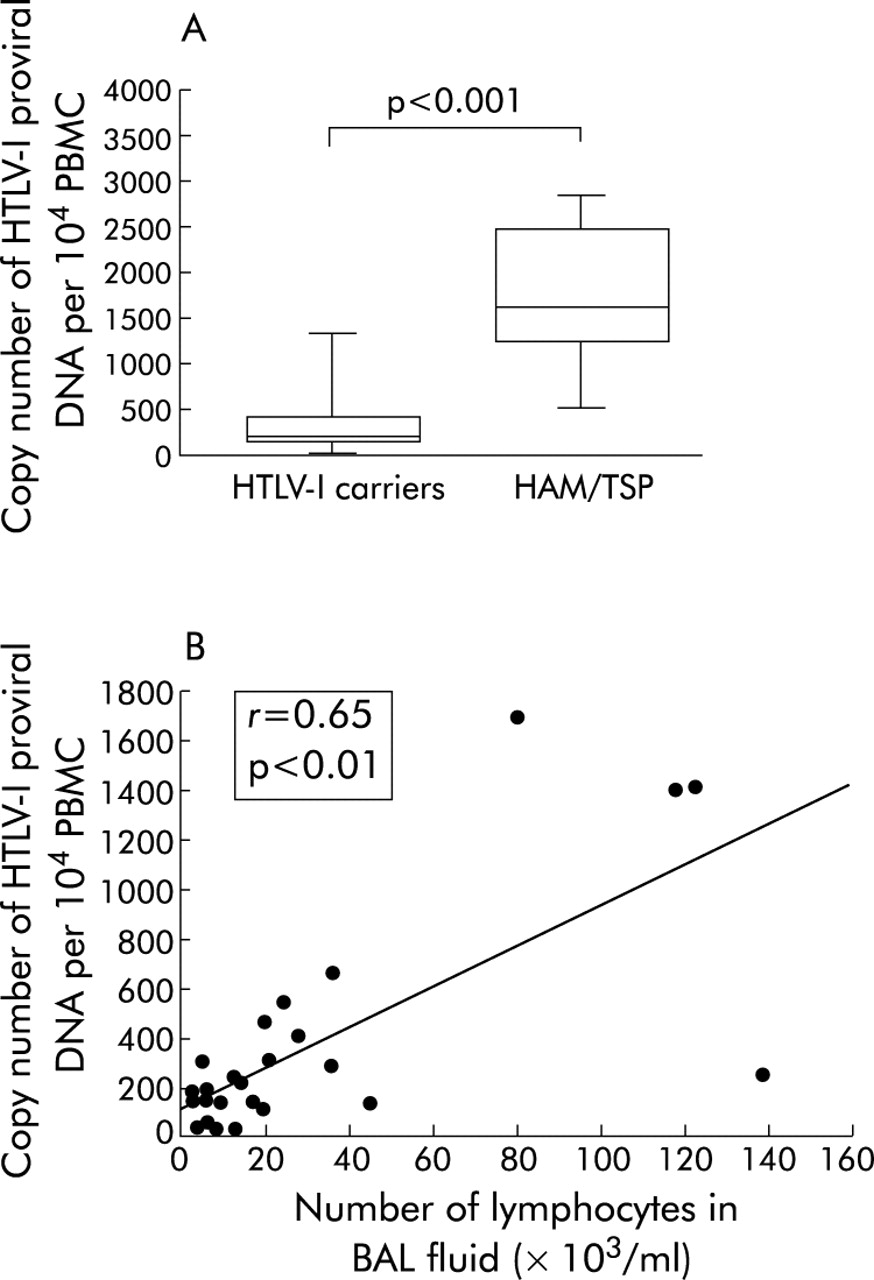
The copy number of HTLV-I proviral DNA in PBMCs was determined in 26 HTLV-I carriers (11 AHCs and 15 SHCs) and eight patients with HAM/TSP. As shown in table 4 and fig 1A, the copy number of HTLV-I proviral DNA per 104 PBMCs was significantly increased in patients with HAM/TSP compared with those of HTLV-I carriers (p<0.001), AHCs (p<0.01), and SHCs (p<0.01). There was no significant difference in the copy number of HTLV-I proviral DNA of PBMCs between AHCs and SHCs.
Quantification of HTLV-I proviral DNA in PBMCs and BAL fluid from HTLV-I carriers and HAM/TSP patients

(A) Copy number of HTLV-I proviral DNA per 104 PBMCs in HTLV-I carriers and patients with HAM/TSP. The whisker box plots represent the 25–75th percentile of results inside the box, the median is shown by the horizontal bar across the box, and whiskers on the box represent the 10–90th percentiles. (B) Correlation between the number of lymphocytes in bronchoalveolar lavage (BAL) fluid and the copy number of HTLV-I proviral DNA per 104 PBMCs in HTLV-I carriers. HTLV-I, human T lymphotropic virus type I; PBMCs, peripheral blood mononuclear cells; HAM/TSP = HTLV-I associated myelopathy/tropical spastic paraparesis.
The relationship between the HTLV-I proviral load in PBMCs and the lymphocyte number in BAL fluid was examined in 26 HTLV-I carriers. As shown in fig 1B, the number of lymphocytes in BAL fluid correlated well with the copy number of HTLV-I proviral DNA in PBMCs of HTLV-I carriers (r = 0.65, p<0.05).
HTLV-I proviral DNA in BAL fluid and correlation with lymphocytes and T lymphocyte subsets in BAL fluid
HTLV-I proviral DNA was detected by PCR in BAL cells from all 10 AHCs examined and was not detected in BAL cells from HTLV-I seronegative controls.
The copy number of HTLV-I proviral DNA in BAL fluid was determined in 14 HTLV-I carriers (three AHCs and 11 SHCs) and in eight patients with HAM/TSP. As shown in table 4, the copy number of HTLV-I proviral DNA per 104 BAL cells was significantly increased in patients with HAM/TSP compared with those of HTLV-I carriers (p<0.001) and SHCs (p<0.01).
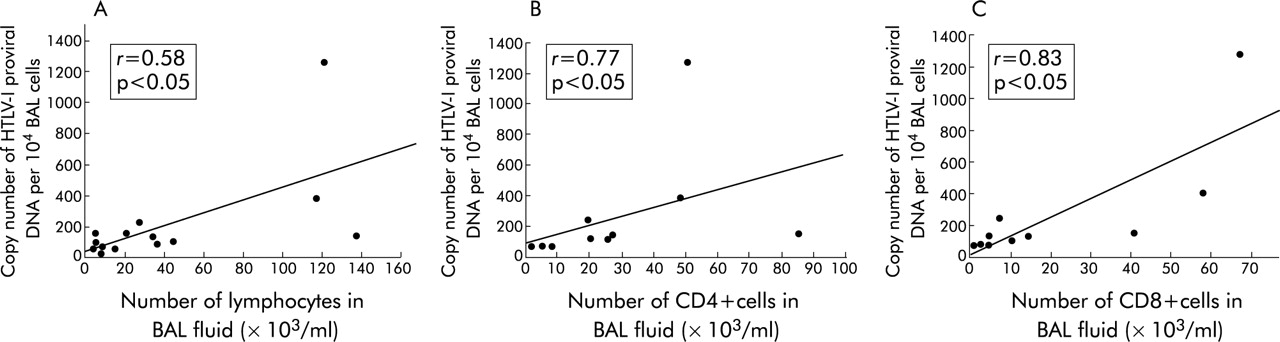
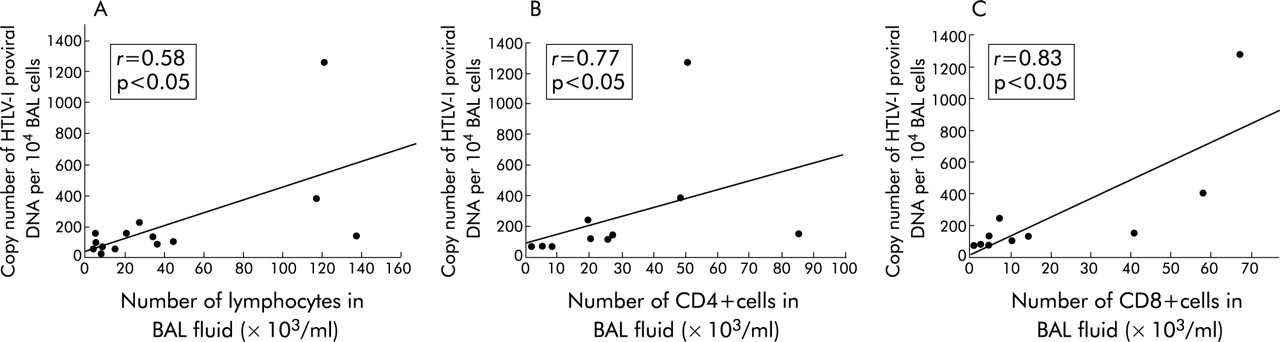
The correlation between HTLV-I proviral DNA load, lymphocyte number, and T cell subsets in BAL fluid was examined in 14 HTLV-I carriers and 10 HTLV-I carriers. The copy number of HTLV-I proviral DNA in BAL fluid correlated well with the number of lymphocytes in BAL fluid from HTLV-I carriers (r = 0.58, p<0.05, fig 2A), CD4+ cells (r = 0.77, p<0.05, fig 2B), and CD8+ cells (r = 0.83, p<0.05, fig 2C).
{kind=link}
{kind=link}
Correlation between (A) the number of lymphocytes in bronchoalveolar lavage (BAL) fluid and the copy number of HTLV-I proviral DNA per 104 BAL cells from HTLV-I carriers; (B) the number of CD4+ cells in BAL fluid and the copy number of HTLV-I proviral DNA per 104 BAL cells from HTLV-I carriers; and (C) the number of CD8+ cells in BAL fluid and the copy number of HTLV-I proviral DNA per 104 BAL cells from HTLV-I carriers. Statistical analyses were performed using Spearman rank correlation. HTLV-I, human T lymphotropic virus type I.
Follow up of asymptomatic HTLV-I carriers with bronchoalveolar lymphocytosis
Three AHCs (subjects 5, 10 and 13) with bronchoalveolar lymphocytosis (>18% lymphocytes in BAL fluid) were available for follow up evaluation of clinical and radiographic variables for 5–10 years. A further individual (subject 9) dropped out 1 year after the study. None of these four individuals showed overt respiratory illness and chest radiographic findings remained normal during the follow up period. They did not develop adult T cell leukaemia, HAM/TSP, or other HTLV-I associated disorders during this time.
DISCUSSION
The major findings of this study are: (1) bronchoalveolar lymphocytosis (>18% lymphocytes in BAL fluid) was observed in 11 of 30 HTLV-I carriers without HAM/TSP or HTLV-I uveitis (36.7%), although there was no significant difference between HTLV-I carriers and control subjects; (2) the number of lymphocytes in the BAL fluid of HTLV-I carriers was significantly correlated with the HTLV-I proviral DNA load in PBMCs; and (3) the HTLV-I proviral DNA load in the BAL fluid was significantly correlated with the number of lymphocytes, CD4+ cells and CD8+ cells in the BAL fluid of HTLV-I carriers.
Recent studies have indicated that immunological dysfunction related to the increased HTLV-I proviral load may be involved in the pathogenesis of HAM/TSP and HTLV-I uveitis.13,16,19–23 However, some studies have shown that these immunological and virological findings are also present in some HTLV-I carriers including relatives of subjects with HAM/TSP and AHCs who have not developed HAM/TSP or HTLV-I uveitis.13,16,20 The genetic background may be implicated in the HTLV-I proviral load and immunological dysfunction in HTLV-I carriers.21,24 These reports suggest that HTLV-I carriers consist of groups of individuals of different genetic backgrounds with various amounts of HTLV-I proviral DNA in PBMCs. Our results showed that the HTLV-I proviral load is high in a subset of HTLV-I carriers without HAM/TSP or HTLV-I uveitis and that the increased HTLV-I proviral load correlates well with bronchoalveolar lymphocytosis in HTLV-I carriers. The present findings, together with those of previous studies, suggest that an increased HTLV-I proviral load may lead to certain systemic conditions including bronchoalveolar lymphocytosis in HTLV-I carriers.
In addition to the genetic background, direct or indirect mechanisms induced by inflammatory conditions may have influenced the HTLV-I proviral load in PBMCs of some SHCs with chronic inflammatory diseases of the respiratory tract in this study, as occurs in HIV-1 infection.25
The HTLV-I proviral load in BAL fluid appeared to be related to the proportion of lymphocytes in the BAL fluid of HTLV-I carriers, as reported previously.26 Interestingly, the HTLV-I proviral load in the BAL fluid correlated with the number of CD8+ cells as well as with the number of CD4+ cells in HTLV-I carriers, even though CD4+ cells are thought to be preferentially infected by HTLV-I.27 Our findings may be consistent with more recent observations of the tropism of HTLV-I to CD8+ lymphocytes.28,29 Further investigations are needed to determine which T cell subsets are predominantly infected with HTLV-I in the lungs of HTLV-I carriers.
A few studies have described some AHCs with bronchoalveolar lymphocytosis who did not develop HAM/TSP and HTLV-I uveitis.30,31 In the present study pulmonary involvement was subclinical in AHCs with bronchoalveolar lymphocytosis, similar to the findings of these previous studies. Furthermore, follow up studies suggested that the clinical development of bronchoalveolar lymphocytosis may be delayed in HTLV-l carriers.
HTLV-I infected lymphocyte(s) commonly exist in the lower respiratory tract of HTLV-I seropositive individuals, as shown in the PCR study of BAL cells. This suggests that factors other than the presence of HTLV-I in the lung—such as a systemic increase in the HTLV-I proviral load, as found in this study—may be necessary for excessive accumulation of lymphocytes in the lung. The mechanisms by which an increased HTLV-I proviral load affects pulmonary involvement in HTLV-I carriers remains to be fully clarified. One possible mechanism is that the increased number of HTLV-I infected cells enhances the probability that infected cells will enter the target organs,31,32 resulting in a local inflammatory response. However, the frequency of HTLV-I specific cytotoxic T lymphocytes22,23,33 related to the increased amount of the virus might be involved in the pathogenesis, as has been shown in lung disorders of patients with HIV infection.25,34
In conclusion, we have shown that pulmonary lymphocytic infiltration can occur in a subset of HTLV-I carriers who have not developed HAM/TSP or HTLV-I uveitis. This pulmonary involvement may be associated with an increased amount of HTLV-I proviral DNA in peripheral blood.