Article Text

Abstract
Background: A number of adhesion molecules are involved in the process of neutrophil infiltration into the lung. P-selectin is one of these neutrophil-endothelial cell adhesion molecules. A study was undertaken to examine the involvement of P-selectin in the development of bleomycin induced inflammation and the ability of N-acetyl-l-cysteine to reduce the potential expression of this selectin in rats.
Methods:N-acetyl-l-cysteine (3 mmol/kg po) was administered daily for seven days prior to bleomycin administration (2.5 U/kg). The kinetics of P-selectin expression and the effect of N-acetyl-l-cysteine after bleomycin treatment were measured using radiolabelled antibodies. P-selectin localisation was evaluated by immunohistochemistry and neutrophil infiltration was assessed by myeloperoxidase activity.
Results: Bleomycin administration resulted in an upregulation of P-selectin at 1 hour, returning to baseline at 3 hours. Myeloperoxidase activity showed a significant increase at 6 hours after bleomycin administration that lasted for 3 days. N-acetyl-l-cysteine treatment completely prevented these increases.
Conclusion: Upregulation of P-selectin in the lung is associated with neutrophil recruitment in response to bleomycin. The beneficial effect of N-acetyl-l-cysteine on bleomycin induced lung injury may be explained in part by the prevention of neutrophil recruitment in the inflammatory stage of the disease.
- bleomycin
- fibrosis
- N-acetyl-l-cysteine
- P-selectin
- inflammation
Statistics from Altmetric.com
Bleomycin is a glycopeptide antibiotic used in the chemotherapy of different types of cancer that produces its antineoplastic effect by causing oxidant damage to DNA.1 Since bleomycin causes an inflammatory process that finally results in pulmonary fibrosis, it is currently used in experimental studies to produce oxidant induced inflammatory and fibrotic lesions in the lung interstitium of various animal species.1,2 This model of pulmonary fibrosis resembles that seen in humans and has been used to assess the effects of potential therapeutic agents.3–5 Bleomycin induced lung fibrosis appears to be the consequence of a primary inflammatory lesion characterised by an accumulation of alveolar macrophages and neutrophils in the lower respiratory tract.6 It has been reported that reactive oxygen species generated from neutrophils can induce lung injury and fibrosis.7
N-acetyl-l-cysteine (NAC) is a mucolytic agent8 that has been well characterised in vitro and in vivo as an agent that protects against oxygen radical damage by its direct scavenger properties.9 NAC is probably one of the most widely investigated drugs and it has been shown to have beneficial effects in clinical conditions in which free radicals are involved.10 NAC instilled intratracheally has been reported to reduce bleomycin induced lung toxicity.5,11 Other authors have reported a significant improvement in lung function after treatment with a high dose of NAC combined with a low dose of steroids in patients with idiopathic pulmonary fibrosis.12
This therapeutic activity of NAC could also be the result of its ability to regulate the production of some cytokines and adhesion molecules in endothelial and bronchial epithelial cells.5 Several authors have reported that the upregulation of different adhesion molecules in various models of lung fibrosis plays a pivotal role in the adhesion of neutrophils to the endothelium during the inflammatory process.13–15
The progression of the inflammatory response involves sequential neutrophil-endothelial cell interactions described as rolling, activation, firm adhesion, and migration.16 These processes are controlled by complex interactions between surface receptors on neutrophils and their corresponding endothelial cell ligands, designated selectins, integrins, or supergene immunoglobulins. Appropriate stimulation of neutrophils or endothelial cells results in an upregulation of the expression of these molecules.16
It is known that E-selectin and intercellular adhesion molecule-1 (ICAM-1) are required for leucocyte migration into inflamed areas17 and both play an important role in bleomycin induced lung injury.14,15 However, the role of P-selectin is not well understood because it shows different patterns of expression in lung injury.18 Several kinds of mediators, including tumour necrosis factor and thrombin, can induce the expression of P-selectin on the endothelial surface within a few minutes.19 Upregulation of P-selectin in endothelial cells can also be triggered by oxygen free radicals but, in this case, P-selectin is steadily expressed for several hours.19
The present study was designed to evaluate the involvement of P-selectin in the development of the inflammatory response caused by bleomycin. Taking into account that bleomycin induced lung injury in rodents is mediated by the production of reactive oxygen species,1,20 we hypothesised that NAC might be able to reduce the potential expression of this selectin. To test this hypothesis we have studied the inflammatory injury and P-selectin expression in bleomycin treated animals, as well as the effect of NAC treatment on this process.
METHODS
Animals
Pathogen free male Sprague-Dawley rats weighing 200–250 g at the start of the experiments were obtained from Criffa (Iffa Credo, France). The rats were housed in a controlled environment and fed rodent chow (A04; Panlab, Barcelona, Spain) and tap water ad libitum. This study conformed to European Community Directive 86/609/EEC for the use of experimental animals and it was approved by the institutional committee of animal care and research.
Chemicals
Bleomycin sulphate was purchased from Almirall-Prodesfarma (Barcelona, Spain), N-acetyl-l-cysteine from Roig Farma (Terrassa, Spain), halothane (Fluothane) from Zeneca Farma (Pontevedra, Spain), sodium pentobarbital from Normon (Madrid, Spain), and thiobutabarbital (Inactin) from Research Biochemicals International (Natick, MA, USA).
Experimental protocol
Lung inflammation and treatments
To produce pulmonary inflammation the animals received endotracheally, by the transoral route, a single dose of 0.25 U bleomycin per 100 g body weight dissolved in 0.25 ml saline (0.9% NaCl). Sham animals received the same volume of intratracheal saline instead of bleomycin. Tracheal instillation was carried out under halothane anaesthesia. For the biochemical assays at each time point (0, 1, 3, 6 and 24 hours, 3 and 15 days) after endotracheal administration of bleomycin or saline, animals were sacrificed by a lethal intraperitoneal injection of sodium pentobarbital (100 mg/kg) followed by exsanguination from the abdominal aorta.
Vehicle (water) or NAC (3 mmol/kg body weight daily; 1 ml final volume) was administered po daily for 7 days before intratracheal instillation of bleomycin (at 09.00 hours).12,21,22
Experimental groups
To determine the kinetics of neutrophil accumulation, animals were randomly distributed into two groups: control (untreated) rats (n=6) and pulmonary inflammation rats (bleomycin, n=6) for each time point (0, 1, 3, 6, 24, and 72 hours). To determine myeloperoxidase (MPO) activity, hydroxyproline levels and histological studies, rats were randomly divided into four groups: sham (vehicle + vehicle, n=6), sham (vehicle + NAC, n=6), bleomycin + vehicle (n=6), and bleomycin + NAC (n=6). Lung tissue samples were obtained, weighed, and processed separately for biochemical (immediately frozen in liquid nitrogen) and histological studies as indicated below.
Additional experiments were performed at the same time points to measure the expression of P-selectin.
Biochemical assays
Myeloperoxidase
Tissue MPO activity, used as an indicator of neutrophil infiltration, was measured spectrophotometrically using 3,3`,5,5`-tetramethylbenzidine as a substrate as previously reported23 with some modifications.24
Hydroxyproline assay
Hydroxyproline levels in lung tissue were used as a quantitative index of fibrogenesis and fibrosis.25 Lung hydroxyproline levels were determined spectrophotometrically as previously reported26 and the results were expressed as μg hydroxyproline per lung.25
Histological study
For histological studies the lung was first perfused through its main bronchus with a fixative solution (10% neutral buffered formalin) at a pressure of 25 cm H2O, immersed in the fixative for 12–24 hours, and blocks were taken. Tissue blocks were placed in formalin, dehydrated in a graded series of ethanol, embedded in paraffin, cut into 4 μm thick serial sections, and stained with haematoxylin-eosin and Masson's trichrome to identify inflammatory cells, connective tissue, and collagen deposition.
Measurement of P-selectin expression
Monoclonal antibodies
Monoclonal antibodies (mAbs) used for the in vivo assessment of P-selectin expression were RMP-1, a murine IgG1 against rat and mouse P-selectin, and P23, a non-binding murine IgG1 directed against human but not rat P-selectin. RMP-1 and P23 were scaled up and purified by protein A/G chromatography at Upjohn Laboratories (Kalamazoo, MI, USA).
Radioiodination of mAbs
Binding mAbs directed against P-selectin were labelled with 125I (Amersham, Madrid, Spain) whereas the isotype matched non-binding mAb P23 was labelled with 131I (Amersham). Radioiodination of mAbs was performed by the iodogen method.27 The specific activity of labelled mAbs was 0.5 mCi/mg.
Experimental protocols
To determine the kinetics of P-selectin expression the right carotid artery and jugular vein were cannulated. After induction of pulmonary inflammation, rats were anaesthetised with an intraperitoneal dose (100 mg/kg) of thiobutabarbital and a mixture of 5 μg 125I-anti-P-selectin mAb (RMP-1), 10 μg unlabelled RMP-1, and 5 μg 131I-non-binding mAb (P23) were administered at 0, 1, 2, and 3 hours. A blood sample was obtained through the carotid artery catheter 5 minutes after injection of the mAbs. The animals were then heparinised (1 mg/kg sodium heparin given intravenously) and rapidly exsanguinated by vascular perfusion of sodium phosphate bicarbonate buffer via the jugular vein, with simultaneous blood withdrawal via the carotid artery, followed by perfusion of sodium phosphate bicarbonate buffer via the carotid artery after the inferior cava vein was severed at the thoracic level. The entire organs were weighed.
The same protocol was used to determine the expression of P-selectin in treated animals.
Calculations
125I (binding mAb) and 131I (non-binding mAb) activities in different organs and in 100 μl aliquots of cell free plasma were counted in a Cobra II gamma counter (Packard, Meridian, Australia). The accumulated activity of each mAb in an organ was expressed as ng binding mAb/g tissue. The formula used to calculate the expression of an adhesion molecule was as follows:
Endothelial expression = ([cpm 125I organ/g/cpm 125I injected] – [cpm 131I organ/g/cpm 131I injected] × [cpm 125I in plasma/cpm 131I in plasma]) × ng injected binding mAb
RNA preparation and RT-PCR of P-selectin
A section of the lungs was processed for total cellular RNA isolation. Total RNA was prepared with TRIzol Reagent following the manufacturer's instructions (GibcoBRL, Life Technologies) and RNA concentrations were calculated from A260 determinations. RNA integrity and loading amounts were assessed by examining 18S and 28S ribosomal RNA banding of samples electrophoresed in 1% agarose gel under non-denaturing conditions and stained with ethidium bromide.
Analysis of P-selectin mRNA expression was made by a semiquantitative RT-PCR method. One μg total RNA was used and the sequences amplified by the Life Technologies one step RT-PCR system according to the manufacturer's protocol. The forward primer was 5`-AGG-TTG-GCA-ATG-GTT-CAC-TC-3` and the reverse primer was 5`-ACC-ATT-GGG-AGC-TAC-ACC-TG-3`. β-actin has been used as a housekeeping gene. In this case, the forward primer was 5`-TCA-TGA-AGT-GTG-ACG-TTG-ACA-TCC-GT-3` and the reverse primer was 5`-CCT-AGA-AGC-ATT-TGC-GGT-GCA-CGA-TG-3`. Sequences were resolved by electrophoresis in denaturing 1.8% agarose gel and stained with ethidium bromide.
Immunohistochemical detection of P-selectin
Primary rabbit polyclonal antibody against rat P-selectin was purchased from Pharmingen (San Diego, CA, USA). Working dilution was 1:50 diluted in 0.05 M Tris buffered saline (TBS), pH 7.6. The secondary biotinylated goat anti-rabbit IgG antibody obtained from Dako (Glostrup, Denmark) was diluted 1:200 in TBS, pH 7.6.
Lung tissue samples on adhesive coated slides were deparaffined, rehydrated, and placed for 30 minutes in a 3% H2O2 solution in distilled water for inhibition of endogenous peroxidase. After washing in TBS (pH 7.6), sections were treated for antigen retrieval. To unmask P-selectin, slides were placed in a 0.1% pronase E solution (Sigma Chemical Co, St Louis, MO, USA) in distilled water at room temperature for 8 minutes. After washing the sections in TBS, endogen biotin was blocked with a commercially available blocking kit (Vector Laboratories Inc, Burlingame, CA, USA). Non-specific binding was blocked with a 10% goat normal serum solution in TBS for 1 hour at room temperature.
The reaction was developed with the avidin-biotin horseradish peroxidase complex (Dako, Glostrup, Denmark) using as chromogen a 0.05% solution of 3,3`-diaminobenzidine (Sigma Chemical Co, St Louis, MO, USA) with 0.03% H2O2 in 0.1 M imidazol buffer (pH 7.1) for 8 minutes. Sections were counterstained with Mayer's haematoxylin. Sections incubated with isotype matched antibodies served as negative controls.
Statistical analysis
Data are expressed as mean (SE) values with 95% confidence intervals (CI) of n experiments. Statistical analysis was carried out by analysis of variance (ANOVA) followed, when differences were significant, by appropriate post hoc tests including the Newman-Keuls test. Differences between the two groups (or between 0 h and different time points in the kinetics analysis) were tested using the Student's t test (GraphPad Software Inc, San Diego, CA, USA). A p value of <0.05 was considered significant.
RESULTS
Myeloperoxidase assay
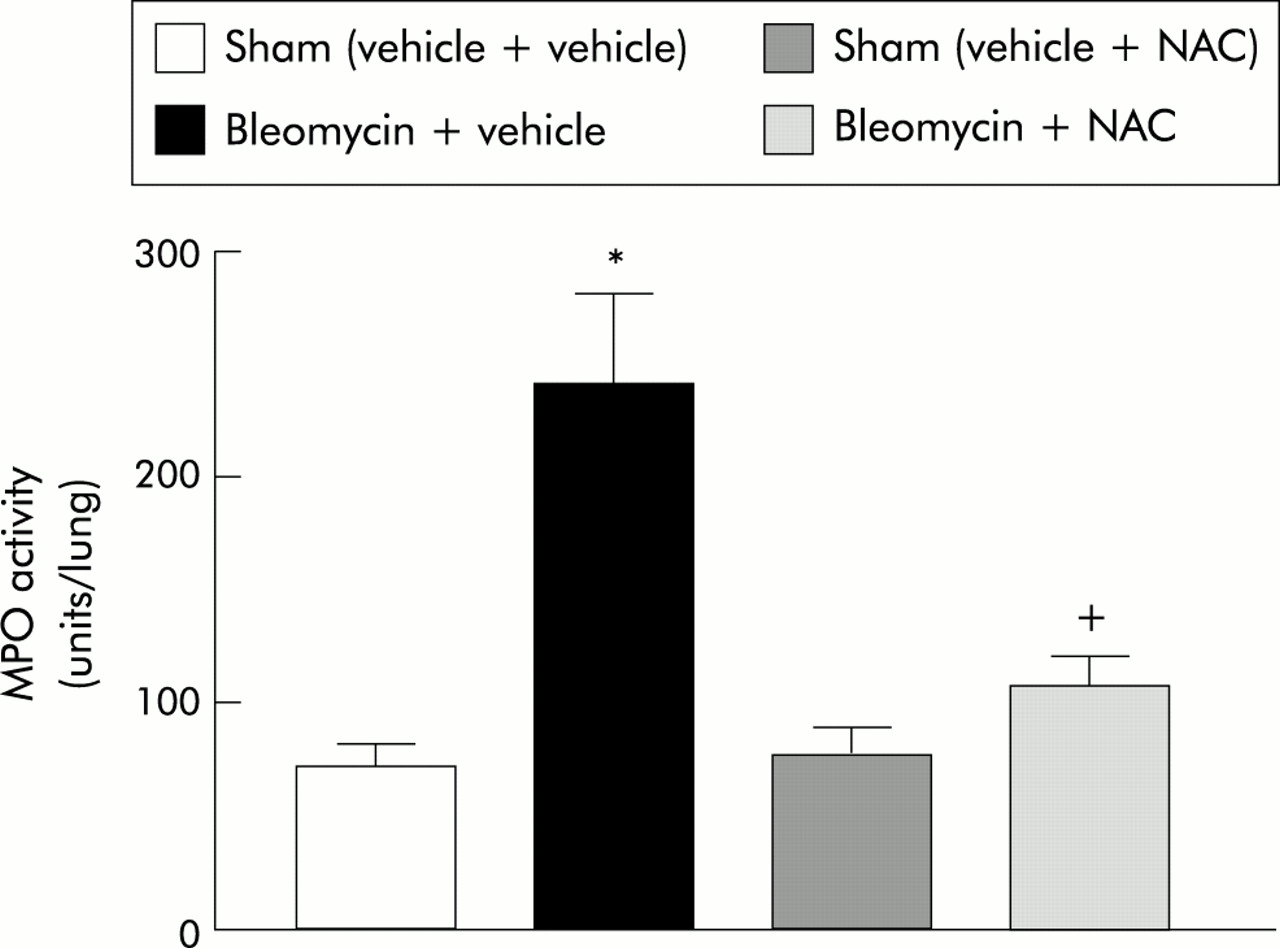
Figure 1 shows the kinetics of MPO activity in lung tissue at 0, 1, 3, 6, 24, and 72 hours after bleomycin administration. An increase in MPO activity was observed 6 hours after the intratracheal instillation of bleomycin (0 h: 64.0 (5.8) units/lung (95% CI 45.6 to 68.8); 1 h: 60.8 (2.6) units/lung (95% CI 54.3 to 67.4); 3 h: 64.5 (3.9) units/lung (95% CI 49.4 to 79.6); 6 h: 98.4 (8.1) units/lung (95% CI 77.7 to 119.1); 24 h: 148.6 (28.0) units/lung (95% CI 78.8 to 218.0); 72 h: 249.7 (24.7) units/lung (95% CI 191.3 to 308.2)).
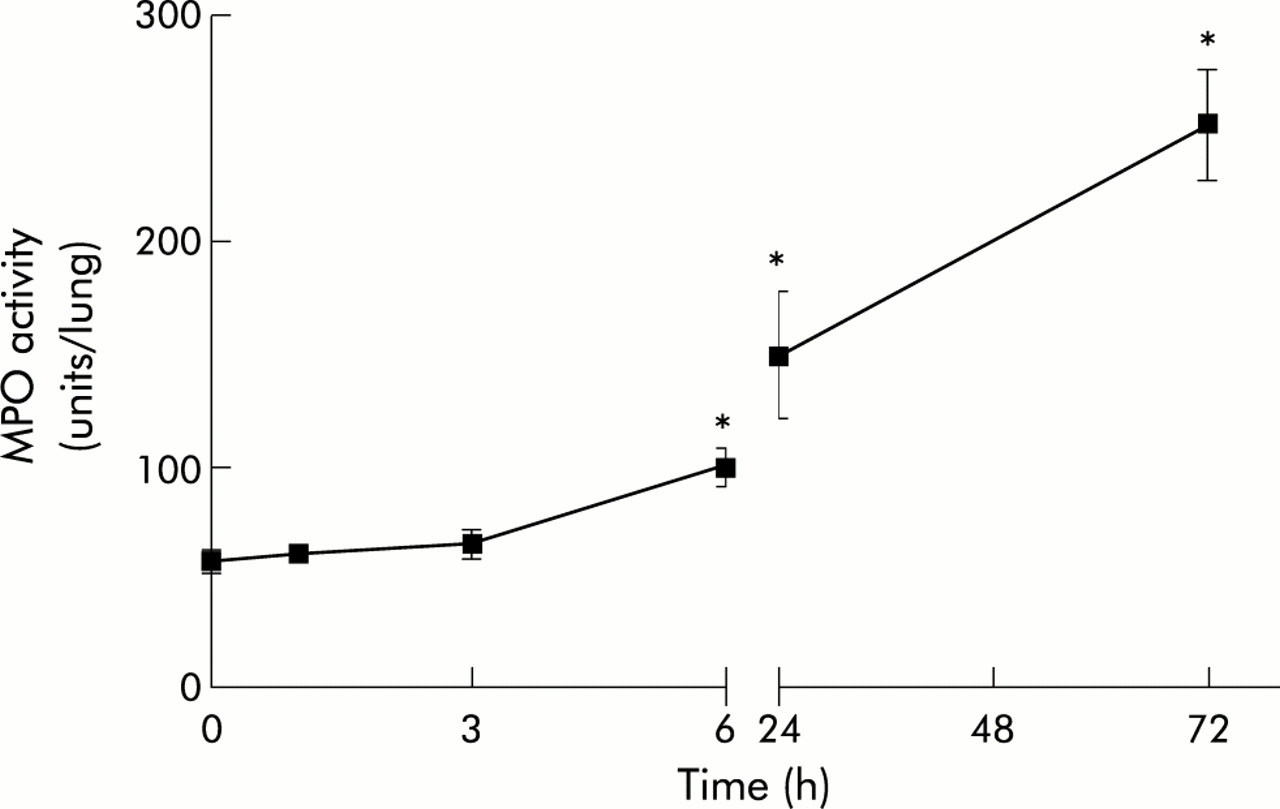
Kinetics of MPO activity in lung tissue after administration of bleomycin (n=6 for each time point). *p<0.05 v 0 h.
Figure 2 shows the effect of NAC on MPO activity in lung tissue 3 days after bleomycin administration. NAC treatment prevented the increase in MPO activity observed in bleomycin treatment (sham (vehicle + vehicle): 70.9 (9.9) units/lung (95% CI 42.9 to 107.5); bleomycin + vehicle: 240.2 (39.4) units/lung (95% CI 164.3 to 320.9); sham (vehicle + NAC): 75.6 (12.3) units/lung (95% CI 27.8 to 128.9); bleomycin + NAC: 106.4 (12.0) units/lung (95% CI 80.5 to 138.2)).

Effect of NAC on MPO activity in lung tissue 3 days after bleomycin administration (n=6 for each group). *p<0.05 v sham groups; +p<0.05 v bleomycin + vehicle.
Hydroxyproline assay
Figure 3 shows hydroxyproline levels 3 days after bleomycin administration (sham (vehicle + vehicle): 2534.0 (137.2) μg/lung (95% CI 2229 to 2840); bleomycin + vehicle: 2930.0 (237.3) μg/lung (95% CI 2383 to 3477); sham (vehicle + NAC): 2314.0 (82.2) μg/lung (95% CI 2052 to 2576); bleomycin + NAC: 2555.0 (250.3) μg/lung (95% CI 2128 to 2983)) and 15 days after bleomycin administration (sham (vehicle + vehicle): 2844.0 (89.1) μg/lung (95% CI 2646 to 3043); bleomycin + vehicle: 6281.0 (384.4) μg/lung (95% CI 4899 to 7663); sham (vehicle + NAC): 2812.0 (79.5) μg/lung (95% CI 2591 to 3032); bleomycin + NAC: 4240.0 (236.2) μg/lung (95% CI 3632 to 4847)). There were no significant differences in any experimental group 3 days after bleomycin administration but 15 days after bleomycin administration there was a significant increase in hydroxyproline levels. Treatment with NAC significantly reduced hydroxyproline levels, although they remained above the level observed in rats not exposed to bleomycin.

Hydroxyproline levels in lung (A) 3 days and (B) 15 days after bleomycin induced lung injury (n=6 for each group). *p<0.05 v sham groups; +p<0.05 v bleomycin + vehicle.
Histological features
Histopathology
Masson trichrome stained lung sections were examined by light microscopy. Lungs from rats in the two sham groups (vehicle + vehicle and vehicle + NAC) were histologically normal (not shown). Lungs from rats in the bleomycin + vehicle group showed a marked increase in interstitial cells with a fibroblastic appearance and deposition of interstitial collagen, and association with focal cuboidal metaplasia of alveolar lining cells (fig 4A). The distribution of lesions was multifocal—that is, patchy areas of pulmonary fibrosis—in most cases, commonly involving the pleura. Although multifocal parenchymal lesions were also present in lungs from rats in the NAC + bleomycin group, there were fewer organised foci and they were smaller than those seen in untreated animals, there was less collagen deposition, and less septal widening (fig 4B).
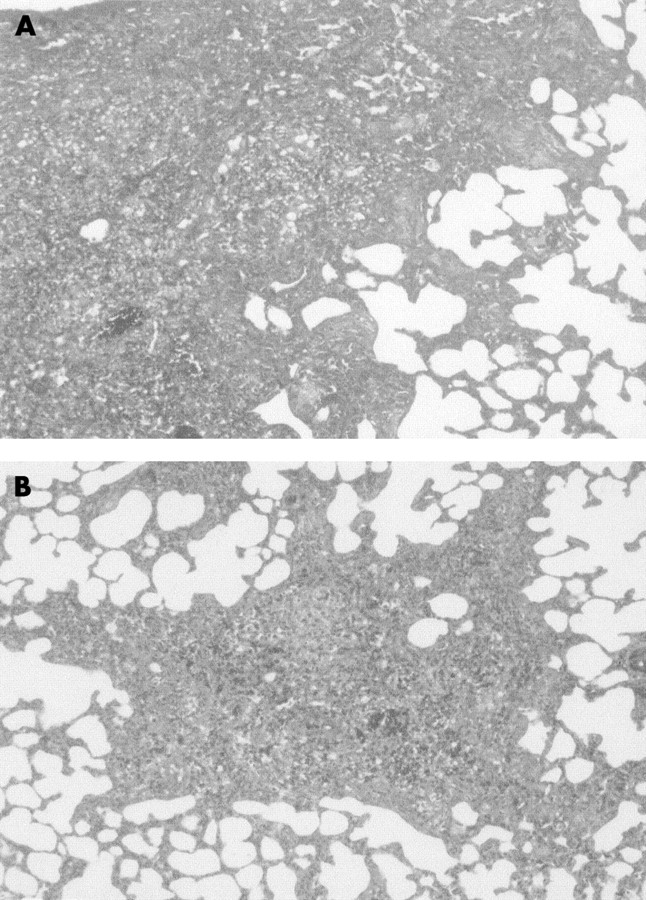
Representative photomicrographs of lung histopathology in (A) bleomycin + vehicle group and (B) bleomycin + NAC group 15 days after endotracheal administration of bleomycin. Lung sections were stained with Masson trichrome. The presence of interstitial collagen (blue staining) was reduced by NAC. Magnification ×250.
P-selectin expression
Figure 5A shows the kinetics of P-selectin expression in the lung. One hour after bleomycin administration P-selectin showed a transient upregulation that returned to baseline levels at 3 hours (0 h: 6.2 (0.5) ng mAb/g (95% CI 4.8 to 7.7); 1 h: 14.0 (1.1) ng mAb/g (95% CI 10.5 to 17.5); 2 h: 7.9 (1.7) ng mAb/g (95% CI 2.5 to 13.3); 3 h: 6.9 (0.7) ng mAb/g (95% CI 4.8–9.0)). Since the maximal expression of P-selectin was at 1 hour, this time point was selected to evaluate the effect of NAC treatment.
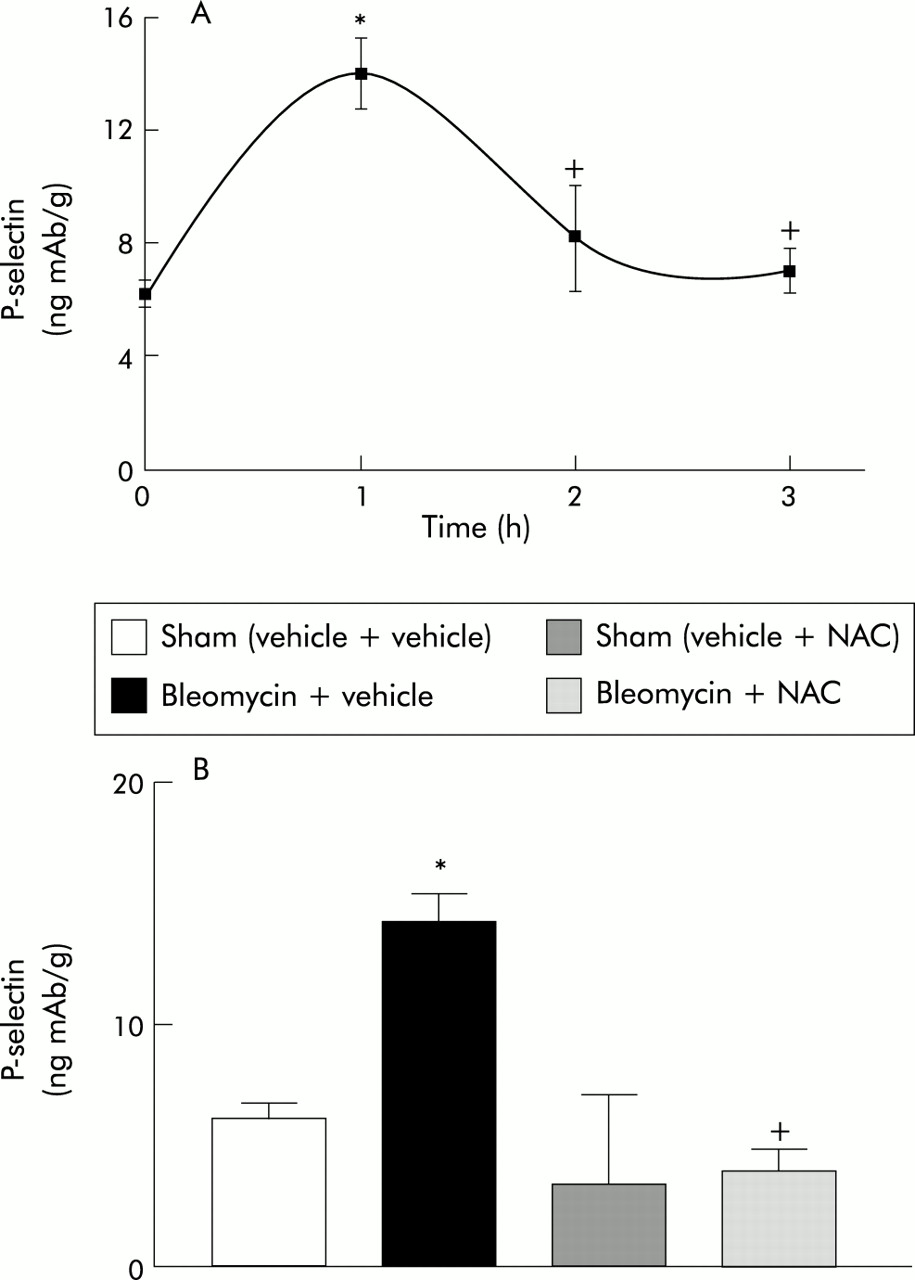
(A) Kinetics of P-selectin expression in lung after bleomycin administration (n=6 for each time point).*p<0.05 v time 0. (B) Effect of NAC treatment on P-selectin expression in the lung 1 hour after bleomycin administration (n=6 for each group). *p<0.05 v sham groups; +p<0.05 v bleomycin + vehicle.
Table 1 summarises the baseline expression of P-selectin in all the organs studied and the effects of bleomycin administration on its expression. P-selectin expression was increased in the lung following administration of bleomycin but it was unchanged or undetectable in the other organs studied.
Levels of P-selectin expression (ng mAb/g) 1 hour after administration of bleomycin
Figure 5B shows the effect of NAC on the expression of P-selectin in lung tissue 1 hour after administration of bleomycin. NAC treatment completely abolished the observed increase in lung P-selectin expression caused by bleomycin (sham (vehicle + vehicle): 6.2 (0.5) ng mAb/g (95% CI 4.8 to 7.7); bleomycin + vehicle: 14.0 (1.1) ng mAb/g (95% CI 10.5 to 17.5); sham (vehicle + NAC): 3.4 (3.2) ng mAb/g (–6.9 to 13.7); bleomycin + NAC: 3.9 (0.7) ng mAb/g (95% CI 1.6 to 6.3)).
RT-PCR of P-selectin
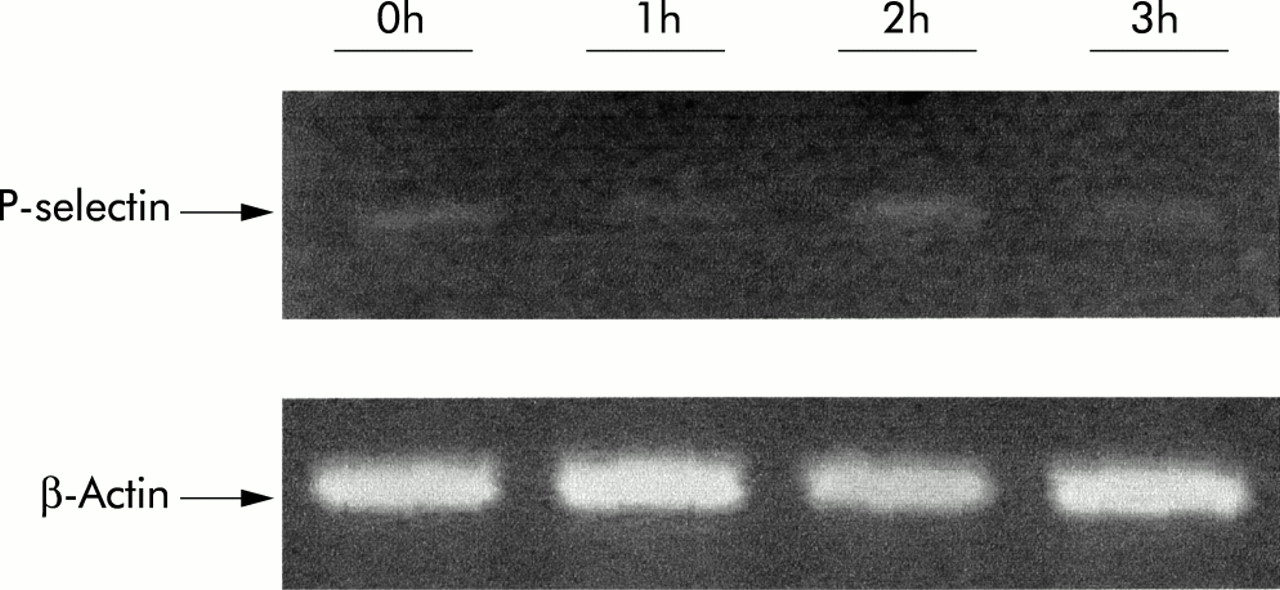
Figure 6 shows the mRNA expression of P-selectin in the lung at 0, 1, 2 and 3 hours, evaluated by RT-PCR. There were no significant differences at any time point after bleomycin administration.

Expression of P-selectin mRNA in lung tissue at different time points after bleomycin administration evaluated by RT-PCR semi-quantitation as described in the text. Amplified sequences were resolved on gel and visualised by ethidium bromide staining. β-actin was used as internal control.
Immunohistochemistry
One hour after administration of bleomycin an intense expression of P-selectin was seen on the endothelial cells of capillaries and small and medium vessels in the bleomycin + vehicle group (fig 7A). By contrast, the expression of P-selectin on the endothelial cells of capillaries and small and medium vessels was moderate in the bleomycin + NAC group (fig 7B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Immunohistochemical analysis of lung parenchyma showing P-selectin (dark staining) on endothelial cells of small blood vessels 1 hour after bleomycin administration. (B) After NAC treatment only a moderate immunostaining on endothelial cells could be observed. Magnification ×100.
DISCUSSION
Two sequential steps appear to be involved in the pathophysiology of bleomycin induced lung injury. The first step is characterised by an early cellular infiltration into the interstitium and alveolar spaces, and the second is the late fibrotic stage. Toxic reactive oxygen species and proteases generated from infiltrated inflammatory cells damage the lung tissue, while excessive fibrosis occurs as a restorative process. Although the mechanism of pulmonary fibrosis in bleomycin treated rats remains unclear, it has been reported that the recruitment of neutrophils plays an important role in the initiation and/or spreading of the fibrogenetic process.28,29 Following neutrophil adhesion to vascular endothelial cells, release of proteases and free radical formation are important processes in cell mediated lung injury, but the role of neutrophils in pulmonary fibrosis is still controversial. It has also been suggested that, in pulmonary fibrosis, increased neutrophils may ameliorate fibrogenesis as a result of an augmented collagenolytic activity and/or decreased collagen synthesis.30
Our results indicate that there is a significant increase in MPO activity, reflecting the neutrophil accumulation observed 6 hours after bleomycin administration and lasting for 3 days (fig 1). This increase could reflect the early inflammatory stage of bleomycin induced lung injury. The late fibrotic process was evidenced by the increase in hydroxyproline levels seen 15 days after bleomycin treatment (fig 3). However, hydroxyproline levels measured 3 days after bleomycin administration showed no significant differences between sham and bleomycin treated animals. These results reflect the biphasic nature of bleomycin induced lung injury.
In previous studies using a similar experimental model5,22 it was observed that NAC exerted an antifibrotic effect. Our results agree with these observations. The increased hydroxyproline levels seen 15 days after bleomycin administration were significantly reduced by NAC pretreatment and histological findings were improved (fig 4A and B). Similarly, it is noteworthy that the increase in MPO activity was also prevented by NAC treatment (fig 1). Some authors have suggested that the late fibrotic process occurs as a consequence of leucocyte mediated cell damage. The effect of NAC on hydroxyproline levels could therefore reflect the inhibition of neutrophil recruitment (fig 1). This hypothesis is supported by previous studies showing that NAC treatment started after the inflammatory stage has no effect on the fibrotic process.5
Since selectins have been implicated in neutrophil mediated acute lung injury,31 we hypothesised that P-selectin upregulation could be involved in the pathogenesis of bleomycin induced lung inflammation. P-selectin is expressed on the surface of endothelial cells and plays an essential role in the initial tethering of neutrophils towards the endothelium, known as rolling.32 Exposure of endothelial cells to oxygen free radicals is known to induce the upregulation of P-selectin.33 An upregulation of P-selectin could be presumed since some of the toxic effects of bleomycin are mediated by free radicals and NAC is a well known free radical scavenger.9,34
When measuring the expression of P-selectin at 1, 2, and 3 hours, our results show that there is an upregulation of P-selectin in the lung 1 hour after the intratracheal instillation of bleomycin that returns to baseline expression at 3 hours (fig 5A). Administration of bleomycin upregulated P-selectin in the lung, but in all other organs studied it remained at baseline levels (table 1). P-selectin is also expressed on platelets but, in our experimental design, animals were exsanguinated and the interference of this expression may be minimal. Immunohistochemical analysis confirmed that the expression of P-selectin was localised in the vascular endothelium (fig 7A, B). These results support the hypothesis that P-selectin expression is one of the events that triggers neutrophil infiltration into the lung during bleomycin induced lung injury.
In a mouse model of bleomycin induced lung injury Azuma et al15 reported a lack of P-selectin mRNA induction. However, as the authors indicated, P-selectin is mainly stored in the Weber-Palade granules and could not be detected by mRNA analysis. Only in other experimental models of lung injury in which expression of P-selectin was maintained35 could mRNA levels be increased. In our model the short period of protein expression in the membrane of endothelial cells was not concomitant with changes in mRNA expression. It could be concluded that this transient expression is related to the mobilisation of the stored pool of P-selectin and not with the genetic induction of protein synthesis.
The mechanism by which NAC limits fibrosis is still uncertain, but it is likely to be related to its ability to reduce the effects of the early inflammatory stage of the disease process. A possible mechanism for reducing the severity of the inflammatory process could be the prevention of P-selectin upregulation which is the first step in the process of neutrophil recruitment. With this in mind, we have examined the ability of NAC to prevent the upregulation of P-selectin in our experimental model. The results show (fig 5B) that NAC treatment effectively prevented P-selectin upregulation 1 hour after bleomycin administration.
In conclusion, our results indicate that P-selectin is upregulated in the early stages of bleomycin induced lung injury and probably plays an important role in neutrophil recruitment. The effect of NAC treatment may be related to its ability to prevent neutrophil recruitment in the inflammatory stages of the disease.
Acknowledgments
This work was supported by grant 1FD97-1143 from the European Union (Regional Development Funds, FEDER) and grant FIS98/1367 (Spanish Ministry of Health).